
カニムシ
| カニムシ | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
| Pseudoscorpiones Haeckel, 1866 | |||||||||||||||
| シノニム | |||||||||||||||
| |||||||||||||||
| 和名 | |||||||||||||||
| カニムシ(蟹虫) |
カニムシ(蟹虫、擬蠍、英語: pseudoscorpion, 学名: Pseudoscorpiones)は、鋏角亜門クモガタ綱に分類される節足動物の分類群の一つ。分類学上はカニムシ目とされる。尾のないサソリに似た姿で、触肢に大きな鋏をもつ[1]。ほとんどは数mm程度以下の微小動物である[2]。世界最大と言われるthe giant pseudoscorpion(Garypus titanius)で体長12−15mm[3]。日本のテナガカニムシは、オスが長大な触肢を持つが、体長は5mmほどである。
特徴
[編集]体は円筒形から楕円形で、途中はくびれがない。全身の外形は、尾のないサソリを彷彿とさせる。4対の歩脚と、最大の特徴である一対の鋏型の触肢をもつ。触肢は大きいものでは体長と同じくらいの長さがあり、先端近くには感覚毛が生えている。この触肢を前方に延ばしてそろそろと歩き、何かにぶつかると、触肢を体に引き付けて、すっ飛ぶように後退する。その姿が印象的なためか、「アトビサリ」等の別名がある。
中には口元の鋏角から糸を出せるものがあり、それを用いて巣を作る。
外部形態
[編集]
o: 側眼、I: 鋏角、II: 触肢基節、III-VI: 歩脚基節、prae-gen: 後体第1節、1-11: 後体第2-12節、an: 肛門
-
 背面
背面 -
 腹面
腹面


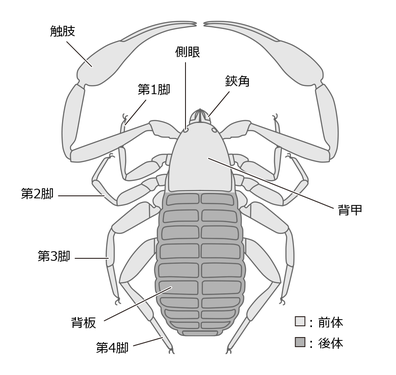
体は大きく前体(prosoma, または頭胸部)と後体(opisthosoma, または腹部)という2つの合体節に分れている。両者の間には、はっきりした境目はあるが、著しいくびれはない。他のクモガタ類と同様、前体は鋏角1対・触肢1対・脚4対という計6対の付属肢(関節肢)をもつ[1]。
前体はおおよそ長方形から三角形の背甲(carapace, prosomal dorsal shield)に覆われる。その前方側面に眼(側眼 lateral eye)がある。眼は単眼で2対もしくは1対、無眼のものもある。腹面は触肢と脚の基部(基節 coxa)によって占められ、左右のそれが正中線で合わさっているため、その間には腹板(sternite)がない[1]。
.jpg)
体の前端には鋏角(chelicera)がある。鋏角は2節の鋏型で[1]、不動指は背面の内側、可動指は腹側の外側にある。大きさは様々で、ツチカニムシ類では頭胸部とほとんど同長なくらいに大きいが、他のものでははるかに小さい。鋏角には紡績腺があるものが多いが、その位置は様々である。
触肢(pedipalp)はよく発達で6節に分れ、本群のとても目立つ特徴となっている。基節は左右癒合して不動だが、転節(trochanter)以降は能動的で左右に広がり、前に折り曲げる。先端2節は大きな鋏(chela)をなし、基部節が掌部と背面の不動指、末端節が腹面の可動指となっている。この鋏には、長い感覚毛があり、これを触角のように用いると同時に、獲物を捕獲するのにも用いる。鋏の両指もしくは片方の指に毒腺があり、捕えた獲物を麻痺させる[4]。また、触肢は種類により性的二形が見られ、例えばヤドリカニムシ科の場合は雄の方が大きい[5]。
鋏角と触肢の間に口が開き、その背面は1枚の目立たない上唇(labrum, 口上板/口上突起 epistome と上唇の複合体 epistomo-labral plate ともされる)、両腹面は触肢基節由来の1対の突起物(lateral lips)に覆われている[6]。
触肢の後方には4対の脚(歩脚 walking leg)が配置する。第1-2脚は前を、第3-4脚は後ろを向く。その構造は第1-2脚、第3-4脚がそれぞれに似ていて、いずれも後方の方がよく発達している。脚の跗節(tarsus)は種類や番目により分節があったりなかったりするため、基節含めて計6節もしくは7節に分かれている[7]。現生種の第3-4脚は、一見してクモガタ類として典型的な短い膝節(patella)をもたないが、古典的に「第1-2腿節」(femur 1-2, basi-telofemur)と呼ばれてきた第3-4肢節は、実際にはそれぞれ短縮した腿節と長大に特化した膝節と考えられる[8]。一部の種類の第1-2脚、および基盤的な化石種においても第3-4肢節が典型的な腿節と膝節の構造を保っている[9]。
後体はほぼ卵形で、付属肢はない。12節からなるが、最終体節は腹面に曲がって肛門を囲んでいる[1]。背面と腹面は、それぞれに独立した背板と腹板に覆われるが、それぞれが正中線で左右に分かれることが多い。生殖孔は腹面第2節の生殖口蓋(genital operculum)に開く[1]。カニムシ科のオスは腹面の生殖口内に、一対の羊角包 ram's horn organs という突起を持ち、求愛行動の最中に体外にたびたび伸長させて求愛に用いる[10]。
呼吸系
[編集]
A: 気門周辺(上から第4脚基節・生殖口蓋・後体第3-4腹板、腹板左右2対の粒が気門)、B: 気管
呼吸器は気管系からなり、その開口である2対の気門は、後体第3と第4節の両腹面に開く[1]。
内部形態
[編集]消化系は、口から肛門に至る消化管であるが、その側面に伸びる腸腺が最も大きい。口の内側に咽頭があり、そこから後方に細い食道が続く。その後方で左右に広がる腸腺がある。腸腺はキチン質の裏打ちがあって、左右に分かれて後方に伸び、さらに外側にいくつもの膨らみを持つ。後体の背板の下は、ほぼこの腸腺に占められる。この後方には、細い小腸が伸び、体内で一度折り返して前に伸び、さらにもう一度折り返して、肛門に続く。
中枢神経系は、前体に大きな集中部があり、これは食道の前後で食道上神経塊と食道下神経塊からなる。
排出器としては、基節腺(coxal gland)、腎嚢などがある。
生態
[編集]カニムシは、主として土の中に生息する土壌生物である。やや大柄なものは、石をめくればその裏面にいるのが見られる。小型のものは、野外で採集するのはほとんど不可能で、土壌動物を採集するための装置が必要になる。イソカニムシは、海岸性の大型種で、岩礁海岸の潮上帯で、岩のすき間や割れ目の間に住んでいる。樹上生活を行う種もいる[11]。小笠原諸島固有種で、オスが長大な触肢を持ち顕著な性的二型を示すテナガカニムシは、タコノキの樹皮下に棲む。
鋏が目立つことでもわかるように、捕食性の動物であり、より小型の動物、トビムシなどを餌にしている。土壌動物としては、密度はさほど高くないが、重要な肉食者である。餌を求めて歩くのではなく、鋏状の触肢に獲物が触れるまで待つ。
昆虫などの脚を掴み、またはその身に乗って便乗する習性が知られる。中南米に分布し、立ち枯れの朽木上で生活するCordylochernes scorpioides は、テナガカミキリの腹部に乗って新しい生活場所に移動することで知られる[12]。アカネズミの巣に棲み、ネズミに寄生するマダニを捕食するオオヤドリカニムシは、ネズミの体毛に掴まり、ネズミとともに移動する[13]。
繁殖行動
[編集]
オスからメスへの精子の受け渡しは、精包(spermatophore=精子塊を包んだカプセル)を介した間接的な方法による。土壌中に棲むツチカニムシ科等では、オスが残置した精包を、メスが見つけて精子を取り込むことが知られている。このようなオスとメスが直接出会うことなく精子を受け渡す方法はトビムシなど他の土壌動物でも行われている。一方、カニムシ科、ヤドリカニムシ科などの種では、オスが触肢でメスの触肢を掴んでメスを自分がその場で作った精包に導き精子を取り込ませることが知られ、その様子は求愛ダンスと呼ばれる[14][10]。メスが貯精嚢(spermatheca)を持つ種では、精子は受精されるまでの間保存されるが、持たない種では、すぐに卵と受精される。受精した卵(胚)は、卵塊として腹部下面の生殖口から外部に張り出した育児嚢の中で孵化するまで保持され、母親の卵巣から供給される栄養を含んだ分泌液(ミルク)を吸収して発育する[14][15]。1度の抱卵数は種により異なり、2~40個。子供が第一若虫となって孵化するまでの間、メスは絹を紡いで小部屋を作って中に閉じこもる種もいる[16]。
分類
[編集]多くのクモガタ類と同様、カニムシの真鋏角類における系統的位置ははっきりしない[1]。かつては形態学の類似、例えば背甲と関節した2節の鋏型の鋏角・短い膝節を持たない(短い腿節と長大な膝節をもつ)歩脚・後体第3-4節にある気門・口周辺の構造などに基づいて、カニムシは一般にヒヨケムシに最も近縁で姉妹群と考えられてきた(共に Haplocnemata/Apatella をなす)[6][17]。しかし2010年代以降、これは分子系統解析に否定的とされ、代わりにサソリや四肺類(クモ・ウデムシ・サソリモドキ・ヤイトムシ)を含んだ単系統群(蛛肺類)との類縁関係を示唆される[17]。この場合、カニムシは蛛肺類の姉妹群[18][19]、もしくは蛛肺類に含め、サソリの姉妹群(共に Panscorpiones をなす)とされる[20][21][22][23]。
下位分類
[編集]以下の科が知られる(和名は日本分類学会連合による[24])。
- ツチカニムシ科 Chthoniidae
- Lechytiidae
- オウギツチカニムシ科 Pseudotyrannochthoniidae
- ケブカツチカニムシ科 Tridenchthoniidae
- Feaellidae
- Pseudogarypidae
- Bochicidae
- Gymnobisiidae
- Hyidae
- Ideoroncidae
- コケカニムシ科 Neobisiidae
- Parahyidae
- ツノカニムシ科 Syarinidae
- イソカニムシ科 Garypidae
- Garypinidae
- リクイソカニムシ科 Geogarypidae
- Larcidae
- Menthidae
- サバクカニムシ科 Olpiidae
- ウデカニムシ科 Cheiridiidae
- Pseudochiridiidae
- Sternophoridae
- メクラカニムシ科 Atemnidae
- カニムシ科 Cheliferidae
- ヤドリカニムシ科 Chernetidae
- イボカニムシ科 Withiidae
脚注
[編集]- ^ a b c d e f g h Lamsdell, James C.; Dunlop, Jason A. (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395–418. ISSN 1467-8039.
- ^ 以下、全般的には内田監(1966)、p.61-90.による。
- ^ Giant pseudoscorpion(Garypus titanius) https://www.iucnredlist.org/species/135739408/135745815#assessment-information
- ^ Krämer, Jonas; Pohl, Hans; Predel, Reinhard (2019-04-15). “Venom collection and analysis in the pseudoscorpion Chelifer cancroides (Pseudoscorpiones: Cheliferidae)”. Toxicon 162: 15–23. doi:10.1016/j.toxicon.2019.02.009. ISSN 0041-0101.
- ^ McLean, Callum J.; Garwood, Russell J.; Brassey, Charlotte A. (2018-11-06). “Sexual dimorphism in the Arachnid orders” (英語). PeerJ 6: e5751. doi:10.7717/peerj.5751. ISSN 2167-8359.
- ^ a b Dunlop, Jason A. (2000). “The epistomo-labral plate and lateral lips in solifuges, pseudoscorpions and mites”. Ekologia Bratislava 19: 67-78.
- ^ “Discover Life in America- GSMNP Pseudoscorpions”. 2022年3月23日閲覧。
- ^ SHULTZ, JEFFREY W. (1989-09-01). “Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications”. Zoological Journal of the Linnean Society 97 (1): 1–56. doi:10.1111/j.1096-3642.1989.tb00552.x. ISSN 0024-4082.
- ^ JUDSON, MARK L. I. (2012-02-20). “Reinterpretation of Dracochela deprehendor (Arachnida: Pseudoscorpiones) as a stem-group pseudoscorpion”. Palaeontology 55 (2): 261–283. doi:10.1111/j.1475-4983.2012.01134.x. ISSN 0031-0239.
- ^ a b G. Kirchmair and G. Raspotnig (2021) Mating behavior of Dactylochelifer latreillii latreillii (Pseudoscorpiones: Cheliferidae): A quantitative study. The Journal of Arachnology 49(2):198-204. https://doi.org/10.1636/JoA-S-20-057
- ^ 佐藤英文、「トゲヤドリカニムシの生活史について」『Acta Arachnologica』 1978-1979年 28巻 1号 p.31-37, doi:10.2476/asjaa.28.31, 日本蜘蛛学会
- ^ David W. Zeh and Jeanne A. Zeh (1992) On the Function of Harlequin Beetle-Riding in the Pseudoscorpion, Cordylochernes scorpioides (Pseudoscorpionida: Chernetidae). Journal of Arachnology 20 (1) : 47-51
- ^ Okabe.K. et al (2018) Tick predation by the pseudoscorpion Megachernes ryugadensis (Pseudoscorpiones: Chernetidae), associated with small mammals in Japan. Journal of the acarological society of Japan. 27 (1) : 1-11. DOI: 10.2300/acari.27.1
- ^ a b Weygoldt, P. 1969 The Biology of Pseudoscorpions. Harvard University Press.
- ^ Adaptations for matrotrophy in the female reproductive system in the pseudoscorpion Chelifer cancroides (Chelicerata: Pseudoscorpiones, Cheliferidae)
- ^ 佐藤英文 2021 カニムシ - 森・海岸・本棚にひそむ未知の虫 築地書館
- ^ a b Dunlop, Jason; Garwood, Russell J. (2014-11-13). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359.
- ^ Sharma, Prashant P.; Fernández, Rosa; Esposito, Lauren A.; González-Santillán, Edmundo; Monod, Lionel (2015-04-07). “Phylogenomic resolution of scorpions reveals multilevel discordance with morphological phylogenetic signal”. Proceedings. Biological Sciences 282 (1804): 20142953. doi:10.1098/rspb.2014.2953. ISSN 1471-2954. PMC 4375871. PMID 25716788.
- ^ Lozano-Fernandez, Jesus; Tanner, Alastair R.; Giacomelli, Mattia; Carton, Robert; Vinther, Jakob; Edgecombe, Gregory D.; Pisani, Davide (2019-05-24). “Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida” (英語). Nature Communications 10 (1): 2295. doi:10.1038/s41467-019-10244-7. ISSN 2041-1723.
- ^ Sharma, Prashant P.; Baker, Caitlin M.; Cosgrove, Julia G.; Johnson, Joanne E.; Oberski, Jill T.; Raven, Robert J.; Harvey, Mark S.; Boyer, Sarah L. et al. (2018-05-01). “A revised dated phylogeny of scorpions: Phylogenomic support for ancient divergence of the temperate Gondwanan family Bothriuridae” (英語). Molecular Phylogenetics and Evolution 122: 37–45. doi:10.1016/j.ympev.2018.01.003. ISSN 1055-7903.
- ^ Benavides, Ligia R.; Cosgrove, Julia G.; Harvey, Mark S.; Giribet, Gonzalo (2019-10-01). “Phylogenomic interrogation resolves the backbone of the Pseudoscorpiones tree of life” (英語). Molecular Phylogenetics and Evolution 139: 106509. doi:10.1016/j.ympev.2019.05.023. ISSN 1055-7903.
- ^ Howard, Richard J.; Puttick, Mark N.; Edgecombe, Gregory D.; Lozano-Fernandez, Jesus (2020-11-01). “Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata” (英語). Arthropod Structure & Development 59: 100997. doi:10.1016/j.asd.2020.100997. ISSN 1467-8039.
- ^ Ontano, Andrew Z; Gainett, Guilherme; Aharon, Shlomi; Ballesteros, Jesús A; Benavides, Ligia R; Corbett, Kevin F; Gavish-Regev, Efrat; Harvey, Mark S et al. (2021-06-01). “Taxonomic Sampling and Rare Genomic Changes Overcome Long-Branch Attraction in the Phylogenetic Placement of Pseudoscorpions”. Molecular Biology and Evolution 38 (6): 2446–2467. doi:10.1093/molbev/msab038. ISSN 1537-1719. PMC 8136511. PMID 33565584.
- ^ “カニムシ目 Pseudoscorpionida - 日本産生物種数調査 - 日本分類学会連合”. www.ujssb.org. 2019年9月3日閲覧。
参考文献
[編集]- 内田亨監修 『動物系統分類学』第7巻(中A)「真正蜘蛛類」、中山書店。
関連項目
[編集]この項目は、動物に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(Portal:生き物と自然/プロジェクト:生物)。 |
| 典拠管理データベース: 国立図書館 |
|---|