
カリョシントリプス
| カリョシントリプス | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 カリョシントリプスの前部付属肢
| |||||||||||||||||||||
| 保全状況評価 | |||||||||||||||||||||
| 絶滅(化石) | |||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||
| 古生代カンブリア紀第四期終盤/ウリューアン期 - ドラミアン期(約5億1,000万 - 5億200万年前) | |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Caryosyntrips Daley & Budd, 2010 [2] | |||||||||||||||||||||
| タイプ種 | |||||||||||||||||||||
| Caryosyntrips serratus Daley & Budd, 2010 [2] | |||||||||||||||||||||
| 種 | |||||||||||||||||||||
カリョシントリプス(Caryosyntrips)は、約5億年前のカンブリア紀に生息したラディオドンタ類と思われる節足動物の一属。くるみ割りに似た前部付属肢のみによって知られ、主に北アメリカから複数の種の化石が発見される[1]。
名称
[編集]学名「Caryosyntrips」は、ギリシア語で種実類を意味する「karyon」と神話での粉砕者の名前「syntrips」の合成語であり、くるみ割りのような形の前部付属肢に因んで名付けられた[2]。
化石と分布
[編集]
カリョシントリプスはほぼ単離した前部付属肢の化石標本のみによって知られている[1][2][3]。最初はカナダブリティッシュコロンビア州の堆積累層バージェス頁岩(バージェス動物群、ウリューアン期、約5億1,000万 - 5億500万年前[4])に産出する化石標本のみ知られていた[2] が、後にアメリカユタ州の堆積累層 Wheeler Formation、Spence Shale、および Marjum Formation(ドラミアン期、約5億200万年前)からにも本属の化石標本が見つかり、一部の種は分布域が複数の堆積累層に及ぶことも判明した[1][3]。また、葉足動物と解釈され、ムレロポディア(Mureropodia)として命名された化石標本 MPZ 2009/1241 の正体は本属の前部付属肢であれば(後述参照)、本属の分布域はスペインの Valdemiedes Formation(Murero biota、カンブリア紀第四期終盤[5]、約5億900万年前)にまで及ぶとなる[1]。
形態
[編集]-
 カリョシントリプスの前部付属肢
カリョシントリプスの前部付属肢 -
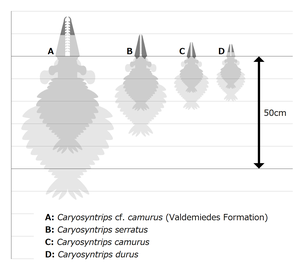 カリョシントリプスのサイズ比較図
カリョシントリプスのサイズ比較図

前部付属肢(frontal appendage)は長い直角三角形のように先端ほど細くなり、常に真っ直ぐに伸びていて、他のラディオドンタ類のように大きく湾曲することはない[2]。唯一に対で保存された前部付属肢の化石標本 ROM 59501 によると、この1対の前部付属肢はハサミのように左右から噛み合せた構造をなしている[2]。各肢節の境目(節間膜)は種や位置によって不明瞭になることはあるが、14節(柄部1節と残り13節[6]、記載当初では12節と誤解釈された[2])が確認されており、最初と最後の肢節を除いてそれぞれの内側には、単調な棘に似た1対の短い内突起(endite)をもつ[1]。C. camurus 以外の種では、前部付属肢の外側にはノコギリ状の小さな棘(dorsal spine, outer spine)が並んでいる[1]。元々頭部に連結したと思われる前部付属肢の基部は、釣り鐘のような形をした突出部をもつ[1]。また、化石標本での各肢節の境目は曲線であったことにより、前部付属肢は元々円錐状であったと考えられる[2]。
上記の化石標本 ROM 59501 は、1対の前部付属肢の付け根に接続した有機物の塊や、少し離れた所に散在した断片が見られており、同一個体の頭部(おそらく甲皮[3])由来の構造であった可能性がある[2]。
体は発見されていないが、前部付属肢の長さをラディオドンタ類であるアノマロカリスとインノヴァティオカリス[7]の体の比率(それぞれの体長は柄部を除いた前部付属肢長の約2倍と2.8倍)にあわせて推算すると、カリョシントリプスの体長は種によって十数cmから30cmにも及ぶとなる(もし MPZ 2009/1241 は本属の前部付属肢であれば、体長は最大50cm以上にも及ぶ)[8]。
生態
[編集]
カリョシントリプスは捕食者であったと考えられる[1][2]。基部の釣り鐘状の突出部を蝶番状の支点とし、左右に動ける頑丈な前部付属肢を節足動物の大顎ないし顎基のように用いて、獲物を捕獲したり食物を切断したりできたと思われる[1][2][6]。この特徴は、上下の湾曲動作によって獲物を捕獲する多くの肉食性ラディオドンタ類、例えばアノマロカリスの前部付属肢とは大きく異なる[1][2]。ラディオドンタ類の中で前部付属肢がこのような機能をもつと考えられるのは、本属の他にスタンレイカリスなど数少ない例のみ挙げられる[6]。
分類
[編集]
| ||||||||||||||||||||||||
| カリョシントリプスの系統的位置[6][8][9][10][11][12] 青枠:ラディオドンタ類 |

カリョシントリプスは前部付属肢が他のラディオドンタ類より特異で、他の部分も不明であるため、ラディオドンタ類としての本質は懐疑的とされる[13][14]。系統解析では、カリョシントリプスはラディオドンタ類の既存のどの科にも属さず、ラディオドンタ類の全ての科よりも早期に分岐した基盤的な群とされている[6][8][9][10][11][12][15][6][16]。なお、カリョシントリプスが(ラディオドンタ類以外の節足動物に対して)他のラディオドンタ類と単系統群になるかどうかなはっきりしておらず、これは系統解析において本属の情報が少なさすぎるのが原因だと考えらる[6]。ラディオドンタ類としての本質が懐疑的なククメリクルスも、似たような解析結果を得られている[11][12]。



カリョシントリプス(カリョシントリプス属 Caryosyntrips)は記載当初の Daley & Budd 2010 では1種(Caryosyntrips serratus)のみと考えられた[2] が、Pates & Daley 2017 以降では少なくとも次の通りに3種(後述の C. cf. camurus を本属の独立種と考えれば4種)を含むことが分かり、前部付属肢の形態によって区別される[1]。
- Caryosyntrips serratus Daley & Budd, 2010[2]
- 本属の模式種(タイプ種)。カナダブリティッシュコロンビア州のバージェス頁岩[2] とアメリカユタ州の Wheeler Shale [1] から発見される。前部付属肢は他の2種より細く、最終肢節には比較的単純な爪をもつ。各肢節の境目は内側で明瞭に見られ、内突起は前方に傾いている。前部付属肢長は最大10.8cm(推算される体長は20-30cm程度)[8]。
- 種小名はラテン語の「ノコギリ状の縁」を意味する「serratus」による[2]。
- Caryosyntrips camurus Pates & Daley, 2017[1]
- カナダブリティッシュコロンビア州のバージェス頁岩、およびアメリカユタ州の Spence Shale [1] と Marjum Formation [3] から発見される。前部付属肢は外側の棘を欠けている。各肢節の境目は外側のみに見られ、先端3節で急に細くなり、最終肢節の爪は長く鈎のように強く湾曲する。内突起はやや後方に傾いている。前部付属肢長は最大7.2cm(推算される体長は14-20cm程度)[8]。
- 種小名はラテン語の「鈎状の」および「湾曲した」を意味する「camurus」による[1]。本種由来の一部の化石標本はかつては C. serratus 由来と解釈された[2]。
- Caryosyntrips durus Pates & Daley, 2017[1]
- アメリカユタ州の Wheeler Shale [1] のみから発見される。前部付属肢は他の2種より太く、先端9節の肢節のみが発見される。各肢節の境目は不明瞭、可動域は上記の2種より低かったと考えられる。先端の肢節は棘や爪を持たず、その直前6つの肢節の外側にはそれぞれ1本の発達した棘をもつ。内突起は比較的に丈夫で短い。完全の前部付属肢長は最大6.4cmと想定される(推算される体長は12-18cm程度)[8]。
- 種小名はラテン語の「硬い」および「曲げられない」を意味する「durus」による[1]。
MPZ 2009/1241に関する議論
[編集]
スペインの堆積累層 Valdemiedes Formation(Murero biota)から発見された化石標本 MPZ 2009/1241 は、Gámez et al. 2011 に葉足動物の前半身と考えられ、新属(ムレロポディア Mureropodia)新種の葉足動物 Mureropodia apae として命名された[5]。しかしこの見解は Stephen & Daley 2017 によって否定され始め、MPZ 2009/1241 は本属由来の前部付属肢の先端6節と見直され(葉足=内突起、頭部と吻=前部付属肢の先端)、本属の新種の可能性をもつ Caryosyntrips cf. camurus として再記載された[1]。
一方で、Gámez et al. 2011 の著者をも含んだ Gámez et al. 2017 は、Stephen & Daley 2017 に対して反論を挙げた。本文献は Gámez et al. 2011 通りに MPZ 2009/1241 を葉足動物と見なし、Stephen & Daley 2017 は、MPZ 2009/1241 の多くの葉足動物的形質、例えば筋組織・触角・突出した吻・鉤爪らしき痕跡を無視していると批判した。同時にカリョシントリプスの特異性(ラディオドンタ類にしては尋常でない節間膜とされる部位の幅狭さや左右に噛み合わせた保存状態)を取り上げて、それに基づいてカリョシントリプスはラディオドンタ類ではなく、むしろカイメン、ヒオリテスやChancelloriidae科などの他の動物であるとの説も提唱した[18]。
この反発に対して、Stephen & Daley 2017 の著者を含んだ Pates et al. 2018 は幾つかの証拠を補足し、Gámez et al. 2017 の見解を否定した。Gámez et al. 2011 と Gámez et al. 2017 に筋組織や触角と思われた痕跡は化石の周りの石基までにも伸びており、該当化石に属しない非生物的な痕跡であることを示した。吻と思われた前部付属肢の先端は葉足動物の吻との類似性が低く、しかも突出した吻は葉足動物の中でオニコディクティオンなどごく一部の種類のみから知られ、葉足動物において一般的な特徴ではない。他の脚の短い葉足動物に比べても、"体"(=前部付属肢の肢節部分)の直径と"葉足"(内突起)の長さの比率は特異的すぎる。"葉足"(内突起)の先にある爪と思われた痕跡も化石の処理過程による痕跡で、葉足動物の鉤爪として一般的な炭素由来の濃い色すら持たない。こうした MPZ 2009/1241 の特徴の多くが葉足動物的ではないと指摘した。同時に、カリョシントリプスはラディオドンタ類であるとこを支持する証拠(カリョシントリプスとカイメン/ヒオリテス/Chancelloriidae類の類似性は大まかな輪郭に過ぎないこと、幅狭い節間膜と左右に噛み合わせた保存状態は、カリョシントリプス以外の少なからぬラディオドンタ類の前部付属肢にも見られることなど)をもいくつか追記した[17]。
脚注
[編集]- ^ a b c d e f g h i j k l m n o p q r s t u v w Stephen Pates; Allison C. Daley (2017). “Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada”. Papers in Palaeontology 3 (3): 461–470. doi:10.1002/spp2.1084.
- ^ a b c d e f g h i j k l m n o p q r s Allison C. Daley, Graham E. Budd (2010). “New anomalocaridid appendages from the Burgess Shale, Canada”. Palaeontology 53 (4): 721–738. doi:10.1111/j.1475-4983.2010.00955.x.
- ^ a b c d e Pates, Stephen; Lerosey-Aubril, Rudy; Daley, Allison C.; Kier, Carlo; Bonino, Enrico; Ortega-Hernández, Javier (2021-01-19). “The diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian: Drumian)” (英語). PeerJ 9: e10509. doi:10.7717/peerj.10509. ISSN 2167-8359.
- ^ Canada, Royal Ontario Museum and Parks (2011年6月10日). “The Burgess Shale” (英語). burgess-shale.rom.on.ca. 2021年7月2日閲覧。
- ^ a b c Gámez Vintaned, José Antonio、Liñán, Eladio、Yu. Zhuravlev, Andrey 著、Pontarotti, Pierre 編(英語)『Evolutionary Biology – Concepts, Biodiversity, Macroevolution and Genome Evolution』Springer Berlin Heidelberg、Berlin, Heidelberg、2011年、193–219頁。doi:10.1007/978-3-642-20763-1_12。ISBN 978-3-642-20762-4。
- ^ a b c d e f g Moysiuk, Joseph; Caron, Jean-Bernard (2021-05). “Exceptional multifunctionality in the feeding apparatus of a mid-Cambrian radiodont” (英語). Paleobiology: 1–21. doi:10.1017/pab.2021.19. ISSN 0094-8373.
- ^ Zeng, Han; Zhao, Fangchen; Zhu, Maoyan (2022-09-07). “Innovatiocaris, a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta” (英語). Journal of the Geological Society 180 (1). doi:10.1144/jgs2021-164. ISSN 0016-7649.
- ^ a b c d e f Lerosey-Aubril, Rudy; Pates, Stephen (2018-09-14). “New suspension-feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton” (英語). Nature Communications 9 (1): 3774. doi:10.1038/s41467-018-06229-7. ISSN 2041-1723.
- ^ a b Vinther, Jakob; Stein, Martin; Longrich, Nicholas R.; Harper, David A. T. (2014-03). “A suspension-feeding anomalocarid from the Early Cambrian” (英語). Nature 507 (7493): 496–499. doi:10.1038/nature13010. ISSN 0028-0836.
- ^ a b Cong, Peiyun; Ma, Xiaoya; Hou, Xianguang; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (2014-09). “Brain structure resolves the segmental affinity of anomalocaridid appendages” (英語). Nature 513 (7519): 538–542. doi:10.1038/nature13486. ISSN 1476-4687.
- ^ a b c Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2015-03-11). “Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps” (英語). Nature 522 (7554): 77–80. doi:10.1038/nature14256. ISSN 0028-0836.
- ^ a b c Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A; Shu, Degan; Paterson, John R (2018-11-01). “Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan”. National Science Review 5 (6): 863–869. doi:10.1093/nsr/nwy057. ISSN 2095-5138.
- ^ Zeng, Han; Zhao, Fangchen; Zhu, Maoyan (2022-09-07). “Innovatiocaris , a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta”. Journal of the Geological Society. doi:10.1144/jgs2021-164. ISSN 0016-7649.
- ^ Potin, Gaëtan J.-M.; Daley, Allison C. (2023). “The significance of Anomalocaris and other Radiodonta for understanding paleoecology and evolution during the Cambrian explosion”. Frontiers in Earth Science 11. doi:10.3389/feart.2023.1160285. ISSN 2296-6463.
- ^ Moysiuk, J.; Caron, J.-B. (2019-08-14). “A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources”. Proceedings of the Royal Society B: Biological Sciences 286 (1908): 20191079. doi:10.1098/rspb.2019.1079. PMC 6710600. PMID 31362637.
- ^ Caron, J.-B.; Moysiuk, J. (2021-09-08). “A giant nektobenthic radiodont from the Burgess Shale and the significance of hurdiid carapace diversity”. Royal Society Open Science 8 (9): 210664. doi:10.1098/rsos.210664.
- ^ a b Pates, Stephen; Daley, Allison; Ortega-Hernández, Javier (2018). “Response to Comment on “Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris” with the formal description of Stanleycaris”. Acta Palaeontologica Polonica 63. doi:10.4202/app.00443.2017.
- ^ Gámez Vintaned, José; Zhuravlev, Andrey (2018-02-01). “Comment on “Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris” by Stephen Pates, Allison C. Daley, and Javier Ortega-Hernández”. Acta Palaeontologica Polonica 63. doi:10.4202/app.00335.2017.
関連項目
[編集]外部リンク
[編集]- Caryosyntrips - Fossil Gallery - The Burgess Shale - カリョシントリプスの簡易ガイド(2011年までの見解に基づくため、前部付属肢は12節と復元され、Caryosyntrips camurus の化石標本 ROM 59501 は Caryosyntrips serratus とされた)