
「ティラノサウルス科」の版間の差分
Pterosauromorpha (会話 | 投稿記録) 編集の要約なし |
テンプレートの修正 |
||
| (8人の利用者による、間の8版が非表示) | |||
| 1行目: | 1行目: | ||
{{出典の明記|date=2017年6月}} |
|||
{{生物分類表 |
{{生物分類表 |
||
|名称 = ティラノサウルス科 |
|名称 = ティラノサウルス科 |
||
|fossil_range = [[白亜紀]] |
|fossil_range = [[後期白亜紀]], {{fossil range|80|66}} |
||
|画像= [[ファイル:Tyrannosauridae Diversity2.jpg|250px]] |
|画像= [[ファイル:Tyrannosauridae Diversity2.jpg|250px]] |
||
|画像キャプション = |
|画像キャプション = |
||
|地質時代 = [[白亜紀]] |
|地質時代 = [[後期白亜紀]] |
||
|地質時代2 = |
|地質時代2 = |
||
|省略 = 恐竜上目 |
|省略 = 恐竜上目 |
||
| 15行目: | 14行目: | ||
|科 = '''ティラノサウルス科''' {{sname||Tyrannosauridae}} |
|科 = '''ティラノサウルス科''' {{sname||Tyrannosauridae}} |
||
|学名 = '''Tyrannosauridae'''<br/>{{AUY|[[ヘンリー・フェアフィールド・オズボーン|Osborn]]|1905}} |
|学名 = '''Tyrannosauridae'''<br/>{{AUY|[[ヘンリー・フェアフィールド・オズボーン|Osborn]]|1905}} |
||
|下位分類名 = [[科 (分類学)|亜科]] |
|下位分類名 = [[科 (分類学)|亜科]]、[[属 (分類学)|属]] |
||
|下位分類 = |
|下位分類 = |
||
* [[アウブリソドン]] |
|||
{{Center|'''本文参照'''}} |
|||
* {{仮リンク|デイノドン|en|Deinodon}} |
|||
* [[アルバートサウルス亜科]] |
|||
* [[ティラノサウルス亜科]] |
|||
}} |
}} |
||
'''ティラノサウルス科''' ({{sname||Tyrannosauridae}}) は、[[ティラノサウルス上科]]に属する[[獣脚類]]の[[恐竜]]の[[科 (分類学)|科]]。同上科の恐竜の中では大型から超大型であり、[[後期白亜紀]]の[[北アメリカ大陸]]と[[アジア]]に生息した。解剖学的特徴は共通性が高いが、幼体やより小型の属種の体は華奢で、大型の属種の成体は頑強な体格を有する<ref name=ポール2020>{{Cite book|和書|title=グレゴリー・ポール恐竜事典 原著第2版 |page=116-117 |author=[[グレゴリー・ポール]] |translator=[[東洋一]]、今井拓哉、河部壮一郎、柴田正輝、関谷透、服部創紀 |date=2020-08-30 |pubisher=[[共立出版]] |isbn=978-4-320-04738-9}}</ref>。 |
|||
[[ティラノサウルス上科|より小型の祖先]]の子孫であるにも拘わらず、ティラノサウルス科の恐竜はほぼ常時各々の[[生態系]]において最大の[[捕食|捕食動物]]であり、[[食物連鎖]]の[[頂点捕食者|頂点]]に位置していた。最大の種 ''Tyrannosaurus rex'' で、これは既知の陸上捕食動物では最大かつ最も頑強であり、全長は12.3メートルを超え<ref name=Hutchinsonetal2011>{{cite journal|last=Hutchinson|first=John R.|author2=Bates, Karl T. |author3=Molnar, Julia |author4=Allen, Vivian |author5=Makovicky, Peter J. |author6= Claessens, Leon |title=A Computational Analysis of Limb and Body Dimensions in ''Tyrannosaurus rex'' with Implications for Locomotion, Ontogeny, and Growth |journal=PLOS ONE |year=2011 |volume=6 |issue=10 |pages=e26037 |doi=10.1371/journal.pone.0026037 |pmid=22022500 |pmc=3192160 |bibcode=2011PLoSO...626037H |doi-access=free}}</ref>、体重は8.4 - 14トンと推定されている<ref name="Hutchinsonetal2011"/><ref name=TH07>{{cite journal |last1=Therrien |first1=F. |last2=Henderson |first2=D. M. |date=2007 |title=My theropod is bigger than yours ... or not: estimating body size from skull length in theropods |journal=Journal of Vertebrate Paleontology |issn=0272-4634 |doi=10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 |volume=27 |issue=1 |pages=108–115|s2cid=86025320 }}</ref><ref name="HartmanMassEstimate">{{cite web |last=Hartman |first=Scott |date=July 7, 2013 |title=Mass estimates: North vs South redux |publisher=Scott Hartman's Skeletal Drawing.com |url=http://www.skeletaldrawing.com/home/mass-estimates-north-vs-south-redux772013 |accessdate=August 24, 2013 |archivedate=October 12, 2013 |archiveurl=https://web.archive.org/web/20131012065922/http://www.skeletaldrawing.com/home/mass-estimates-north-vs-south-redux772013 }}</ref>。二足歩行性であり、顎にはD字型の断面を持つ歯がU字型に配列し、吻部の先端は丸みを帯びる。長い後肢からは他の大型獣脚類と比較して移動速度の向上が示唆されるが、その一方で前肢は手としての機能を保持しつつも退化しており、指も退縮して2本になっている<ref name=ポール2020/>。 |
|||
'''ティラノサウルス科''' ({{sname||Tyrannosauridae}}) は、[[ティラノサウルス上科]]に属する[[恐竜]]の一群である。 |
|||
== |
== 発見の歴史 == |
||
[[Image:Deinodon.JPG|thumb|left|''Deinodon'' の歯。最初期に知られたティラノサウルス科の化石]] |
|||
どこか基板的な[[コエルロサウルス類]]や遠縁の[[カルノサウルス類]]を連想させがちな[[プロケラトサウルス科]](例→[[グアンロン]])とは違い、ティラノサウルス科は、頭は大きく歯も頑丈。前肢が貧弱な代わりに後肢は長く、そして胴体は頑強で身体も大きいと、一般的にイメージされる“ティラノサウルスの仲間”を体現したような身体付きをしていた。 |
|||
ティラノサウルス科の最初の化石が発見されたのは{{仮リンク|カナダ地質調査所|en|Geological Survey of Canada}}による遠征の際であり、その際には無数の離散した歯が発見された。これらの明らかな恐竜の歯は1856年に[[ジョセフ・ライディ]]が ''[[デイノドン|Deinodon]]''(「怖ろしい歯」)として命名した。ティラノサウルス科の最初の良好な標本は[[アルバータ州]]の[[ホースシューキャニオン累層]]で発見されたものであり、部分的な骨格を伴うほぼ完全な頭蓋骨から構成されていた。これらの化石は1876年に[[エドワード・ドリンカー・コープ]]が最初に研究し、東方のティラノサウルス上科の属[[ドリプトサウルス]]の種のものであると考えた。1905年、[[ヘンリー・フェアフィールド・オズボーン]]はアルバータ州の化石をドリプトサウルスと顕著に異なるものであると認識し、''[[アルバートサウルス|Albertosaurus sarcophagus]]''(「肉を食べるアルバータのトカゲ」)と命名した<ref name=carpenter1992/>。1892年にコープは他のティラノサウルス類の骨を記載しており、記載された単離した椎骨は''[[マノスポンディルス|Manospondylus gigas]]'' と命名された。この発見は100年以上に亘って見落とされており、やがて本標本が実際には''[[ティラノサウルス|Tyrannosaurus rex]]'' のものであり、さらに当該の学名に対して先取権を持つことが判明すると、2000年代初頭に議論を呼んだ<ref name=quinlanetal2007b>{{cite conference |first1=B.H. |last1=Breithaupt |last2=Southwell |first2=E.H. |last3=Matthews |first3=N.A. |date=18 October 2005 |title=In Celebration of 100 years of ''Tyrannosaurus rex'': ''Manospondylus gigas'', ''Ornithomimus grandis'', and ''Dynamosaurus imperiosus'', the Earliest Discoveries of ''Tyrannosaurus rex'' in the West |conference=2005 Salt Lake City Annual Meeting |conference-url=http://gsa.confex.com/gsa/2005AM/finalprogram/index.html |book-title=Abstracts with Programs |volume=37 |issue=7 |publisher=Geological Society of America |pages=406 |url=http://gsa.confex.com/gsa/2005AM/finalprogram/abstract_96150.htm |accessdate=8 October 2008 |archivedate=22 October 2019 |archiveurl=https://web.archive.org/web/20191022084528/https://gsa.confex.com/gsa/2005AM/finalprogram/abstract_96150.htm }}</ref>。 |
|||
[[Image:Aublysodon.jpg|thumb|[[アウブリソドン]]に割り当てられた歯]] |
|||
1905年にアルバートサウルスを命名する論文を発表したオズボーンは、同年にさらなるティラノサウルス類の2標本 を記載した。これらの標本は[[アメリカ自然史博物館]]が1902年に行った[[バーナム・ブラウン]]率いる遠征において[[モンタナ州]]と[[ワイオミング州]]で発見されたものであり、オズボーンは当初これらの標本を別種のものであると考えた。彼は1つの標本に ''Dynamosaurus imperiosus''(「力のトカゲの皇帝」)、もう1つの標本に ''Tyrannosaurus rex''(「暴君トカゲの王」)と命名した。1年後、オズボーンはこれらの2標本が同じ種に由来するものであると認識した。最初に発見されたのはディナモサウルスの方であったが、ティラノサウルスという名前が原記載において1ページ先に記載されていたため、ティラノサウルスの名前が用いられるようになった<ref name="Breithaup">{{Cite journal|last=Breithaup|first=BH|author2=Southwell EH |author3=Matthews NA |year=2006|title=''Dynamosaurus imperiosus'' and the earliest discoveries of ''Tyrannosaurus rex'' in Wyoming and the West|journal=New Mexico Museum of Natural History and Science Bulletin |volume=35 |pages=257–258}}</ref>。 |
|||
バーナム・ブラウンはアルバータ州からさらなるティラノサウルス科標本を蒐集しており、当該グループに特徴的な2本指で短い前肢を保存したもの(1914年に[[ローレンス・ラム]]が ''[[ゴルゴサウルス|Gorgosaurus libratus]]'' 「均衡のとれた熾烈なトカゲ」として命名)もその中には含まれていた。ゴルゴサウルスに関わる2つ目の重大な発見は1942年のもので、保存状態が良好であるが異様に小型の完全な頭蓋骨が発見された。当該標本は[[第二次世界大戦]]後に[[チャールズ・W・ギルモア]]が研究し、''Gorgosaurus lancesnis'' と命名した<ref name=carpenter1992/>。1988年には[[ロバート・T・バッカー]]と[[フィリップ・J・カリー]]およびマイケル・ウィリアムズがこの頭蓋骨を再研究し、新属 ''[[ナノティラヌス|Nanotyrannus]]'' に分類した<ref name="bakkeretal1988">{{cite journal |author1=Bakker R.T |author2=Williams M. |author3=Currie P. | year = 1988 | title = ''Nanotyrannus'', a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana | journal = Hunteria | volume = 1 | pages = 1–30 }}</ref>。また、1946年に[[ソビエト連邦]]の古生物学者たちは[[モンゴル国]]への遠征を開始し、アジア初のティラノサウルス類の化石を発見した。1955年に Evgeny Maleev はモンゴルの化石をティラノサウルスおよびゴルゴサウルス、そして1つの新属 ''[[タルボサウルス|Tarbosaurus]]''(「怖ろしいトカゲ」)と命名した。後の研究で、Maleev のティラノサウルス類の種は実際には成長段階の異なるタルボサウルスの種であることが判明した。後にモンゴルで発見された第二の種は1976年にセルゲイ・クルザノフが記載・命名した ''[[アリオラムス|Alioramus remotus]]''(「隔たった異なる枝」)であった。ただし、アリオラムスが真のティラノサウルス科であるか、あるいはより基盤的なティラノサウルス類であるかについては、いまだ議論がある<ref name="kurzanov1976"/><ref name=carpenter1992/>。 |
|||
=== 頭部 === |
|||
上顎の前方(前上顎骨)にあるD字型の断面をした歯、および癒合した鼻骨が共通の特徴である。 |
|||
== 説明 == |
|||
[[File:Tyrannosauridae Size Diagram by PaleoGeek.svg|thumb|left|[[ティラノサウルス]]、[[タルボサウルス]]、[[アルバートサウルス]]、[[ゴルゴサウルス]]、[[ダスプレトサウルス]]の大きさ比較]] |
|||
コエルロサウルス類であることから[[羽毛]]を持っていた可能性が指摘されていたが、[[2004年]]にティラノサウルス科の[[ディロング]]の発見により証明された。さらに比較的大型のティラノサウルス類である[[ユウティラヌス]]にも羽毛の痕跡が見つかり、本科の大型恐竜にも羽毛が生えている可能性が深まった。 |
|||
ティラノサウルス科はいずれも大型の動物であり、全ての種が体重1トンを超える<ref name=holtz2004>{{Cite book|last=Holtz |first=Thomas R. |year=2004 |chapter=Tyrannosauroidea |editor= Weishampel, David B. |editor2=Dodson, Peter |editor3=Osmólska, Halszka |title=The Dinosauria |edition=Second |publisher=University of California Press |location=Berkeley |pages=111–136 |isbn=978-0-520-24209-8}}</ref>。[[アリオラムス]]の1個の標本は全長5 - 6メートルの個体と推定されるものも発見されているが<ref name=kurzanov1976>{{Cite journal|last=Kurzanov |first=Sergei M. |title=A new carnosaur from the Late Cretaceous of Nogon-Tsav, Mongolia |language=ru |journal=The Joint Soviet-Mongolian Paleontological Expedition Transactions |volume=3 |pages=93–104}}</ref>、これを未成熟個体と考える専門家もいる<ref name=holtz2004/><ref name=currie2003a>{{Cite journal |last=Currie |first=Philip J. |year=2003 |title=Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta |journal=Acta Palaeontologica Polonica |volume=48 |issue=2 |pages=191–226 |url=http://www.app.pan.pl/archive/published/app48/app48-191.pdf |accessdate=2010-06-12 |archivedate=2018-08-19 |archiveurl=https://web.archive.org/web/20180819050324/http://www.app.pan.pl/archive/published/app48/app48-191.pdf }}</ref>。[[アルバートサウルス]]、[[ゴルゴサウルス]]、[[ダスプレトサウルス]]はいずれも全長8 - 10メートルであり<ref name=russell1970>{{Cite journal|last=Russell |first=Dale A. |year=1970 |title=Tyrannosaurs from the Late Cretaceous of western Canada |journal=National Museum of Natural Sciences Publications in Paleontology |volume=1 |pages=1–34}}</ref>、[[タルボサウルス]]は12メートルに達する<ref name=maleev1955b>{{Cite journal |last=Maleev |first=Evgeny A. |year=1955 |title=New carnivorous dinosaurs from the Upper Cretaceous of Mongolia |journal=Doklady Akademii Nauk SSSR |volume=104 |issue=5 |pages=779–783 |language=ru |url=http://home.comcast.net/~theropod-archives/pdf/Maleev_1955_DokAkadNauk_104(5)_779_NOC_T.pdf |accessdate=2008-09-10 |archivedate=2012-11-03 |archiveurl=https://web.archive.org/web/20121103131744/http://home.comcast.net/~theropod-archives/pdf/Maleev_1955_DokAkadNauk_104(5)_779_NOC_T.pdf }}</ref>。ティラノサウルスの最大級の標本の一つである[[スー (ティラノサウルス) |スー]]は全長12.3メートルに達する<ref name=Hutchinsonetal2011/>。 |
|||
一方他のティラノサウルス科の恐竜(アルバートサウルス、ダスプレトサウルス、ゴルゴサウルス、タルボサウルス)を調査した結果、それらが鱗状の皮膚で覆われていた可能性が高いと指摘する研究結果もある。それによると、ティラノサウルス科の恐竜が大型化の過程で二次的に羽毛を失ったのではないかとする。ただし保温の難しい幼体についてはなお羽毛を持っていたとする見解もある。 |
|||
=== |
=== 頭蓋骨と歯列 === |
||
{{multiple image |
|||
ティラノサウルスや[[アルバートサウルス]]など科の後期の属では2本の指を持つ非常に短い腕が特徴的である。ティラノサウルスの上腕骨は、大腿骨の3分の1の長さまで退縮していた{{Sfn|犬塚則久|2006|p=140}}。しかしディロングや[[エオティラヌス]]などティラノサウルス上科の初期群は、他のコエルロサウルス類と同じように3本指の長い腕を持つため、これらの特徴はグループ内で2次的に発達した特長であることが分かっている。腕の縮小化は[[ケラトサウルス類]] {{sname||Ceratosauria}} の[[アベリサウルス科]] {{sname||Abelisauridae}} でも体の大型化に伴って起きており関連が注目される。これは、大型化とともに頭骨が巨大になり、重量バランス上、前肢が短縮した方が生存に有利であったためと推定されている{{Sfn|犬塚則久|2006|p=139}}。 |
|||
| align = right |
|||
| total_width = 320 |
|||
|perrow = 2 |
|||
| image1 = Lythronax.png |
|||
=== 後肢 === |
|||
| caption1 = [[リトロナクス]] |
|||
ティラノサウルスやアルバートサウルスに代表されるティラノサウルス科は、中足骨の中央が両隣から挟まれた[[アークトメタターサル]]構造(arctometatarsus)と呼ばれる特殊な構造の足をしていた。この構造は走る際に足へかかる負荷を分散しやすくする効果があり、大抵は俊足の恐竜に見られる構造である。例えば[[オルニトミムス科]]{{sname||Ornitomimidae}}や[[トロオドン科]] {{sname||Troodontidae}}がその好例で、とりわけオルニトミムス科とティラノサウルス科は[[収斂進化]]ないし[[軍拡競争]]の結果アークトメタターサル構造を獲得したものと見られている<ref>MANEUVERED OUT OF A CORNER: LIGAMENT ENTHESES OF THE ARCTOMETATARSUS ENHANCED TYRANNOSAURID AGILITY</ref>。 |
|||
| image2 = Nanuqsaurus hoglundi.png |
|||
ちなみに前述のティラノサウルス科やオルニトミムス科などは、共通してアークトメタターサル構造を持つことから共に[[アークトメタターサリア]] (Arctometatarsalia) と呼ばれるグループを成すと考えられたが、現在では初期の種では見られない特徴であるため[[収斂進化|収斂]]であるとされている。 |
|||
| caption2 = [[ナヌークサウルス]] |
|||
| image3 = Skull and jaws of the holotype (MOR 590) of Daspletosaurus horneri.jpg |
|||
== 分類上の位置 == |
|||
| caption3 = [[ダスプレトサウルス]] |
|||
旧い分類ではTyrannosauridaeは、ジュラ紀の[[アロサウルス]]・[[ケラトサウルス]]など共に旧来の食肉竜類([[カルノサウルス類]] {{sname||Carnosauria}})に分類されていた。しかし近年の[[分岐分類学]]的な研究によれば、頭部や[[坐骨]]の構造などから小型獣脚類のグループである[[コエルロサウルス類]] {{sname||Coelurosauria}} に含まれるとされる。つまり大型のティラノサウルスは、アロサウルスなどのジュラ紀の大型獣脚類の子孫ではなく小型獣脚類が進化で巨大化したものである。白亜紀後期の序盤を過ぎた頃、アロサウルス類が姿を消したのに伴い、小型獣脚類だったティラノサウルス類の一部は大型化への道をたどっていった。 |
|||
| image4 = Teratophoneus.png |
|||
* [[有羊膜類]] {{sname|en|Amniote|Amniota}} |
|||
| caption4 = [[テラトフォネウス]] |
|||
** [[竜弓類]] {{sname||Sauropsida}} |
|||
}} |
|||
*** [[爬虫類]] {{sname|en|Reptile|Reptilia}} |
|||
部分的な頭蓋骨だけが発見されているアリオラムスを除いて全ての属で完全な頭蓋骨が得られているため、ティラノサウルス科の頭蓋骨の解剖学はよく理解されている<ref>{{Cite book|last=Currie |first=Philip J. |year=2000 |chapter=Theropods from the Cretaceous of Mongolia |title=The Age of Dinosaurs in Russia and Mongolia |location=Cambridge |publisher=Cambridge University Press |pages=434–455 |isbn=978-0-521-54582-2}}</ref>。ティラノサウルス、タルボサウルス、ダスプレトサウルスの頭蓋骨は長さ1メートルを超える<ref name="russell1970"/>。成体のティラノサウルス科は高く頑強な頭蓋骨を持ち、数多くの骨が癒合して強度を増している。同時に、数多くの頭蓋骨の間に存在する空洞のチャンバーや、それらの骨の間に位置する大型の開口部(窓)は、頭蓋骨の軽量化に寄与した。高い[[前上顎骨]]や癒合した[[鼻骨]]など、ティラノサウルス科の頭蓋骨に見られる数多くの特徴は直近の祖先にも見ることが出来る<ref name=holtz2004/>。 |
|||
**** [[双弓類]] {{sname|en|Diapsid|Diapsida}} |
|||
***** [[主竜類|主竜形類]] {{sname||Archosauromorpha}} |
|||
****** [[主竜類]] {{sname|en|Archosaur|Archosaurs}} |
|||
******* [[恐竜]] {{sname|en|Dinosaur|Dinosauria}} |
|||
******** [[竜盤類]] {{sname||Saurischia}} |
|||
********* [[獣脚類]] {{sname||Theropoda}} |
|||
********** [[テタヌラ類]] {{sname||Tetanurae}} |
|||
*********** [[コエルロサウルス類]] {{sname||Coelurosauria}} |
|||
************ '''ティラノサウルス類''' {{sname||Tyrannosauridae}} |
|||
ティラノサウルス科の頭蓋骨には数多くの固有の形質がある。具体的には、卓越した{{仮リンク|矢状稜|en|Sagittal crest}}を伴う癒合した[[頭頂骨]]が挙げられる。この稜は{{仮リンク|矢状縫合|en|Sagittal suture}}に沿って縦方向に走り、頭蓋天井の2つの上[[側頭窓]]を隔てる。これらの窓の後側では、ティラノサウルス科は特徴的に高い項部のクレストを持ち、このクレストは縦方向よりもむしろ[[横断面]]に沿って頭頂骨上を走る。このクレストはティラノサウルス、タルボサウルス、アリオラムスにおいて発達する。アルバートサウルス、ダスプレトサウルス、ゴルゴサウルスは[[涙骨]]上に目の前に高いクレストを持つが、タルボサウルスとティラノサウルスは極端に厚みを帯びた[[後眼窩骨]]が三日月形のクレストを目の後ろで形成する。アリオラムスはその吻部の最上部に1列に配列した6個の骨質のクレストを持つ。ダスプレトサウルスとタルボサウルスではより低いクレストが報告された標本があり、さらに基盤的なティラノサウルス上科である[[アパラチオサウルス]]とも共通する<ref name=currie2003a/><ref name=carretal2005>{{Cite journal|last=Carr |first=Thomas D. |author2=Williamson, Thomas E. |author3= Schwimmer, David R. |year=2005 |title=A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama |journal=Journal of Vertebrate Paleontology |volume=25 |issue=1 |pages=119–143 |doi=10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2|s2cid=86243316 |issn=0272-4634}}</ref>。吻部と頭蓋骨の他の部位にも無数の孔が存在する。''[[ダスプレトサウルス|D. horneri]]'' を記載した2017年の研究によると、[[ワニ]]やティラノサウルス科に見られる多列の神経血管孔は触覚や鱗状の外皮にも相関がある<ref>{{Cite journal|last1=Carr| first1=Thomas D. |last2=Varricchio |first2=David J.|last3=Sedlmayr |first3=Jayc C. |last4=Roberts |first4=Eric M. |last5=Moore |first5=Jason R. |date=2017-03-30 |title=A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system |journal=Scientific Reports |language=en |volume=7 |issue=1 |page=44942 |doi=10.1038/srep44942 |issn=2045-2322 |pmid=28358353 |bibcode=2017NatSR...744942C |pmc=5372470}}</ref>。 |
|||
=== 主な属 === |
|||
{{multiple image |
|||
* [[アレクトロサウルス]] {{snamei||Alectrosaurus}} |
|||
| align = left |
|||
* [[アウブリソドン]] {{snamei||Aublysodon}} |
|||
| total_width = 220 |
|||
* [[アリオラムス]] {{snamei||Alioramus}} |
|||
|perrow = 1 |
|||
* [[イテミルス]] {{snamei||Itemirus}} |
|||
| image1 = B-rex teeth.jpg |
|||
* [[シャンシャノサウルス]] {{snamei||Shanshanosaurus}} |
|||
| image2 = The Childrens Museum of Indianapolis - Tyrannosaurus rex tooth.jpg |
|||
* [[キアンゾウサウルス]] {{snamei||Qianzhousaurus}} |
|||
| footer = ティラノサウルスの歯 |
|||
* [[リトロナクス]] {{snamei||Lythronax}} |
|||
}} |
|||
* [[テラトフォネウス]] {{snamei||Teratophoneus}} |
|||
ティラノサウルス科はティラノサウルス上科の祖先と同様に[[異歯性]]を持ち、前上顎骨歯は断面がD字型で、他の歯よりも小型である。最初期のティラノサウルス上科や他の獣脚類と異なり、成熟したティラノサウルス科の[[上顎骨]]歯と[[歯骨]]歯はブレード状ではなく極端に厚く、また断面は丸みを帯びており、いくつかの種では鋸歯が減少している<ref name=holtz2004/>。歯の本数は種のうちで一定しており、大型の種では小型の種よりも少ない傾向がある。例えば、アリオラムスは76 - 78本、ティラノサウルスは54 - 60本の歯を持つ<ref name=currieetal2003>{{Cite journal |last=Currie |first=Philip J. |author2=Hurum, Jørn H |author3=Sabath, Karol. |year=2003 |title=Skull structure and evolution in tyrannosaurid phylogeny |journal=Acta Palaeontologica Polonica |volume=48 |issue=2 |pages=227–234 |url=http://www.app.pan.pl/archive/published/app48/app48-227.pdf |accessdate=2010-10-02 |archivedate=2008-10-31 |archiveurl=https://web.archive.org/web/20081031093118/http://www.app.pan.pl/archive/published/app48/app48-227.pdf |url-status=live }}</ref>。 |
|||
* [[ティラノサウルス亜科]] |
|||
** [[ティラノサウルス]] {{snamei||Tyrannosaurus}}([[タイプ (分類学)|模式属]]) |
|||
** [[タルボサウルス]] {{snamei||Tarbosaurus}} |
|||
** [[ズケンティラヌス]]{{snamei||Zhuchengosaurus}} |
|||
** [[ダスプレトサウルス]] {{snamei||Daspletosaurus}} |
|||
** [[ナノティラヌス]] {{snamei||Nanotyrannus}} |
|||
** [[ナヌークサウルス]]{{snamei||Nanuqsaurus}} |
|||
* [[アルバートサウルス亜科]] |
|||
** [[アルバートサウルス]] {{snamei||Albertosaurus}} |
|||
** [[ゴルゴサウルス]] {{snamei||Gorgosaurus}} |
|||
ウィリアム・エイブラーは2001年に、アルバートサウルスの歯の鋸歯が歯の割れ目に類似し、膨大部と呼称される丸い空洞に帰着することを観察した<ref name="abler-2001">Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84–89. In: ''Mesozoic Vertebrate Life''. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.</ref>。ティラノサウルス科の歯は体から肉を引っ張るための留め具として用いられ、そのためティラノサウルス類が肉片を引き剥がす際の張力によって割れ目のような鋸歯が歯の一面に広がった可能性がある<ref name="abler-2001" />。しかし、膨大部があれば、これらの力をより大きな表面積に分散させ、負担のかかった歯が損傷するリスクを軽減できた<ref name="abler-2001" />。空洞に帰着する切れ込みの存在は、人間の工学とも相等しい。エイブラーの説明によると、[[ギター]]メーカーは空洞に帰着する切れ込みを入れることで柔軟性と剛性を交互に木材に与えるとのことである<ref name="abler-2001" />。また[[航空機]]の表面の保護においては、ドリルで膨大部様構造を作り、素材の亀裂の伝播を防ぐ方法が用いられている<ref name="abler-2001" />。エイブラーは切れ込みとドリルによる穿孔を伴う[[アクリル樹脂]]のバーが規則的に切り込みを入れただけのものに比べて25%以上強い強度を持つことを実証した<ref name="abler-2001" />。ティラノサウルス類や他の獣脚類と異なり、[[植竜類]]や[[ディメトロドン]]などの太古の捕食動物は摂食の力を受けても歯の切れ目状の鋸歯が広がらないようにする適応を遂げなかった<ref name="abler-2001" />。 |
|||
<gallery> |
|||
ファイル:Alioramus BW.jpg|アリオラムス想像図 |
|||
ファイル:Tarbosaurus MPC-D 100 60 skull (3).jpg|タルボサウルス頭骨 |
|||
ファイル:Daspletosaurus torosus steveoc.jpg|ダスプレトサウルス生態復元想像図 |
|||
ファイル:Drumheller 150.jpg|ゴルゴサウルス骨格 |
|||
</gallery> |
|||
== |
=== 体骨格 === |
||
[[File:Journal.pone.0079420.g001.tif|thumb|[[リトロナクス]]と[[テラトフォネウス]]の骨格ダイアグラム]] |
|||
以前は[[白亜紀]]後期を代表するグループであったが白亜紀前期からも知られるようになった。 |
|||
頭蓋骨は厚いS字型の頸部の先端に位置しており、長く重厚な尾は頭部と胴部に対する[[カウンターウエイト]]として機能し、[[重心]]は臀部に位置する。ティラノサウルス科はプロポーション的に非常に小型の2本指の前肢で知られているが、[[痕跡器官 (生物)|痕跡器官]]となった第III指が残る場合もある<ref name=holtz2004/><ref name=quinlanetal2007a>{{Cite journal |last=Quinlan |first=Elizabeth D. |author2=Derstler, Kraig |author3=Miller, Mercedes M. |year=2007 |title=Anatomy and function of digit III of the ''Tyrannosaurus rex'' manus |journal=Geological Society of America Annual Meeting — Abstracts with Programs |page=77 |url=http://gsa.confex.com/gsa/2007AM/finalprogram/abstract_132345.htm |accessdate=2008-01-15 |archivedate=2008-02-24 |archiveurl=https://web.archive.org/web/20080224015023/http://gsa.confex.com/gsa/2007AM/finalprogram/abstract_132345.htm |url-status=live }} [abstract only]</ref>。 |
|||
[[File:Gorgosaurus forelimb.png|thumb|150px|left|[[ゴルゴサウルス]]の肩甲骨と右前肢]] |
|||
ティラノサウルス科は後肢で歩行するため、その骨は頑強である。前肢とは対照的に、大半の他のどの獣脚類と比較しても後肢は体サイズに対して長い。幼体やいくつかの小型の成体では、基盤的ティラノサウルス上科と同様に、[[脛骨]]が[[大腿骨]]よりも長く、[[オルニトミモサウルス類]]と同様に脚の速い恐竜の形質を示す。より大型の成体は動作の緩慢な動物の後肢のプロポーション形質を有しているが、[[アベリサウルス科]]や[[カルノサウルス類]]のような他の大型獣脚類に見られるほどではない。ティラノサウルス科の第III[[中足骨]]は第IIおよび第IV中足骨に挟まれて細くなっており、[[アークトメタターサル]]として知られる構造をなす<ref name=holtz2004/>。 |
|||
アークトメタターサルが最初に進化した時代は不明である。[[ディロング]]のような最初期のティラノサウルス上科にアークトメタターサルは存在しないが<ref name=xuetal2004>{{Cite journal|last1= Xu Xing |author2=Norell, Mark A. |author3=Kuang Xuewen |author4=Wang Xiaolin |author5=Zhao Qi |author6= Jia Chengkai. |year=2004 |title=Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids |journal=Nature |volume=431 |issue=7009 |pages=680–684 |doi=10.1038/nature02855 |pmid= 15470426 |first1= X|bibcode=2004Natur.431..680X |s2cid=4381777 |author1-link=Xu Xing (paleontologist) }}</ref>、より新しい時代の[[アパラチオサウルス]]には見られる<ref name=carretal2005/>。この構造は[[トロオドン科]]や[[オルニトミモサウルス類]]および{{仮リンク|カエグナトゥス科|en|Caenagnathidae}}を特徴づけるが<ref name=holtz1994>{{Cite journal|last=Holtz |first=Thomas R. |year=1994 |title=The phylogenetic position of the Tyrannosauridae: implications for theropod systematics |journal=Journal of Paleontology |volume=68 |issue=5 |pages=1100–1117 |jstor=1306180|doi=10.1017/S0022336000026706 |s2cid=129684676 }}</ref>、最初期のティラノサウルス上科に存在しないことから当該の形質が[[収斂進化]]によるものであることが示唆される<ref name=xuetal2004/>。 |
|||
== 生息地域 == |
|||
[[アビアティラニス]]や[[ジュラティラント]]といった基盤的なティラノサウルス上科の恐竜はヨーロッパから産出しており<ref>{{Cite web|url=https://gendai.ismedia.jp/articles/-/78328?page=2 |title=恐竜は過酷な“山越え”をしていた! ジュラ紀に存在した巨大山脈とは? |date=2020-12-17 |accessdate=2021-02-26 |publisher=[[講談社]] |author=[[小林快次]]}}</ref>、ティラノサウルス科を包括するパンティラノサウルス類はヨーロッパからアジア・北アメリカ・南アメリカ・オーストラリアへ分布を広げたと推測されている<ref>{{Cite web|url=https://gendai.ismedia.jp/articles/-/80078?page=3 |title=南半球からも発見! 自らの足で世界中に広がったティラノ軍団! |publisher=講談社 |author=小林快次 |date=2021-02-11 |accessdate=2021-02-26}}</ref>。アジアと北アメリカではパンティラノサウルス類のうちエウティラノサウルス類が進化し<ref>{{Cite web|url=https://gendai.ismedia.jp/articles/-/80078?page=4 |title=南半球からも発見! 自らの足で世界中に広がったティラノ軍団! |publisher=講談社 |author=小林快次 |date=2021-02-11 |accessdate=2021-02-26}}</ref>、北アメリカのパンティラノサウルス類からティラノサウルス科が出現した。 |
|||
== 分類 == |
|||
ティラノサウルス科恐竜の分布の拡大については2つの仮説が提唱されている。1つは2013年に提唱されたもので、北アメリカ大陸西側([[ララミディア大陸]])で進化を遂げたティラノサウルス科は[[海退]]に伴う陸地の拡大と共に分布を広げ、やがてアジアへ渡ったという説である。この説ではティラノサウルス科の移動は北米からアジアへの一度のみとなる。もう1つは2016年に提唱されたもので、前者よりも複雑な移動があったとされている。こちらでは8000万年前までに[[アリオラムス]]と[[キアンゾウサウルス]]に代表されるアリオラムス族が、7350万年前までに他のティラノサウルス科がアジアへ渡り、後に[[ティラノサウルス]]が北米へ戻ったとされる<ref>{{Cite web|url=https://gendai.ismedia.jp/articles/-/79405?page=2 |title=ティラノ軍団の巨大化はどのように起こったのか? 軍団メンバーを一覧で紹介! |date=2021-01-21 |accessdate=2021-02-26 |publisher=[[講談社]] |author=[[小林快次]]}}</ref>。 |
|||
'''Deinodontidae''' という名前は1866年にエドワード・ドリンカー・コープがこの科について作り<ref name=cope1966>{{cite journal | author = Cope E.D. | year = 1866 | title = Discovery of a gigantic dinosaur in the Cretaceous of New Jersey | journal = Proceedings of the Academy of Natural Sciences of Philadelphia | volume = 18 | pages = 275–279 }}</ref>、1960年代まで新たな名前 Tyrannosauridae の立ち位置で用いられた<ref name=maleev1955>{{cite journal | author = Maleev E.A. | year = 1955 | title = New carnivorous dinosaurs from the Upper Cretaceous Mongolia | journal = Doklady Akademii Nauk SSSR | volume = 104 | issue = 5| pages = 779–783 }}</ref>。Deinodontidae のタイプ属は ''[[デイノドン|Deinodon]]'' で、[[モンタナ州]]産の単離した歯から命名された<ref name=leidy1856>{{Cite journal|last=Leidy |first=Joseph |author-link=ジョセフ・ライディ |year=1856 |title=Notice of remains of extinct reptiles and fishes, discovered by Dr. F.V. Hayden in the badlands of the Judith River, Nebraska Territory |journal=Proceedings of the Academy of Natural Sciences of Philadelphia |volume=8 |pages=72–73}}</ref>。しかし、1970年の北アメリカのティラノサウルス類のレビューにおいて、[[デイル・ラッセル]]は ''Deinodon'' が有効な分類群でないと結論し、Deinodontidae の代わりに Tyrannosauridae の名前を用い、[[国際動物命名規約]]と一致すると主張した<ref name=russell1970/>。このため、ティラノサウルス科という名前が現在の専門家から好まれている<ref name=carpenter1992/>。 |
|||
''Tyrannosaurus'' は1905年に[[ヘンリー・フェアフィールド・オズボーン]]が命名し、これに伴ってティラノサウルス科も命名された<ref name=osborn1905>{{Cite journal|last=Osborn |first=Henry F. |author-link=ヘンリー・フェアフィールド・オズボーン |year=1905 |title=''Tyrannosaurus'' and other Cretaceous carnivorous dinosaurs |journal=Bulletin of the American Museum of Natural History |volume=21 |pages=259–265 |doi=10.1111/j.1468-5965.2007.00735_17.x|issue=3|hdl=2246/1464 }}</ref>。この名前は[[古代ギリシア語]]の ''τυραννος'' (''{{lang|grc-Latn|tyrannos}}'')(「暴君」)および {{lang|grc|σαυρος}} (''{{lang|grc-Latn|sauros}}'')(「トカゲ」)に由来する。[[接尾詞]] ''-idae'' は動物の科の名前に一般に付されるものであり、複数名詞を示すギリシア語の接尾詞 ''-ιδαι'' ''-{{lang|grc-Latn|idai}}'' に由来する<ref name=liddellscott>{{Cite book |last=Liddell |first=Henry G. |author2=Scott, Robert |year=1980 |title=Greek-English Lexicon |edition=Abridged |publisher=Oxford University Press |location=Oxford |isbn=978-0-19-910207-5 |url=https://archive.org/details/lexicon00lidd }}</ref>。 |
|||
==脚注== |
|||
{{脚注ヘルプ}} |
|||
{{Reflist}} |
|||
== |
=== 分類法 === |
||
[[File:Tarbosaurus and Tyrannosaurus.jpg|thumb|一般化されたタルボサウルス(A)とティラノサウルス(B)の差異を示すダイアグラム]] |
|||
* {{Cite book|和書 |
|||
ティラノサウルス科は[[リンネ式階層分類体系]]における[[科 (分類学)|科]]であり、[[ティラノサウルス上科]]と[[獣脚亜目]]の内部に置かれる。 |
|||
|author = [[犬塚則久]] |
|||
|title = 恐竜ホネホネ学 |
|||
ティラノサウルス科は2つの亜科に細分される。[[アルバートサウルス亜科]]は北アメリカの属である[[アルバートサウルス]]と[[ゴルゴサウルス]]を含み、[[ティラノサウルス亜科]]は[[ダスプレトサウルス]]、[[テラトフォネウス]]、[[ビスタヒエヴェルソル]]、[[タルボサウルス]]、[[ナヌークサウルス]]、[[ズケンティラヌス]]、[[ティラノサウルス]]を含む<ref name="FT2014">{{Cite journal| last1 = Fiorillo | first1 = A. R. | last2 = Tykoski | first2 = R. S. | year = 2014| title = A Diminutive New Tyrannosaur from the Top of the World| editor-last = Dodson| editor-first = Peter| editor-link = Peter Dodson| journal = [[PLoS ONE]]| volume = 9| issue = 3| pages = e91287| doi = 10.1371/journal.pone.0091287| ref = {{sfnRef|Fiorillo & Tykoski|2014}} | pmid=24621577 | pmc=3951350| bibcode = 2014PLoSO...991287F| doi-access = free }}</ref>。''Gorgosaurus libratus'' をアルバートサウルス属に、''Tarbosaurus bataar'' をティラノサウルス属に含む研究者もいるが<ref name=carretal2005/><ref name=carpenter1992>{{Cite book|last=Carpenter |first=Ken. |year=1992 |chapter=Tyrannosaurids (Dinosauria) of Asia and North America |editor=Mateer, Niall J. |editor2=Chen Peiji |title=Aspects of Nonmarine Cretaceous Geology |location=Beijing |publisher=China Ocean Press |pages=250–268}}</ref><ref name=paul1988>{{Cite book |last=Paul |first=Gregory S. |author-link=グレゴリー・ポール |year=1988 |title=Predatory Dinosaurs of the World |location=New York |publisher=Simon & Schuster |pages=[https://archive.org/details/predatorydinosau00paul/page/464 464pp] |isbn=978-0-671-61946-6 |url=https://archive.org/details/predatorydinosau00paul/page/464 }}</ref>、分かれた属としてゴルゴサウルスとタルボサウルスを維持することを好む研究者もいる<ref name=holtz2004/><ref name=currie2003a/>。アルバートサウルス亜科はティラノサウルス亜科と比較してより細身の体格、低い頭蓋骨、プロポーション的に長い脛骨を特徴に持つ<ref name=holtz2004/>。ティラノサウルス亜科では、頭頂骨の矢状クレストが[[前頭骨]]上に続く<ref name=currie2003a/>。2014年には、Lü Junchang ''et al.'' がティラノサウルス科内の[[族 (分類学)|族]]として[[アリオラムス]]と[[キアンゾウサウルス]]を含むアリオラムス族を記載した。彼らの系統解析では本族はティラノサウルス亜科の基底に置かれることが示唆された<ref name="Junchang2014">{{cite journal |last1=Lü |first1=Junchang |last2=Yi |first2=Laiping |last3=Brusatte |first3=Stephen L. |last4=Yang |first4=Ling |last5=Li |first5=Hua |last6=Chen |first6=Liu |title=A new clade of Asian Late Cretaceous long-snouted tyrannosaurids |journal=Nature Communications |date=2014 |volume=5 |page=3788 |doi=10.1038/ncomms4788|pmid=24807588 |doi-access=free |bibcode=2014NatCo...5.3788L }}</ref><ref>{{cite web|url=http://www.cbc.ca/news/technology/pinocchio-rex-long-snouted-tyrannosaur-discovered-in-asia-1.2634651 |title=Pinocchio rex long-snouted tyrannosaur discovered in Asia |date=7 May 2014|accessdate=8 May 2014 |archivedate=2 February 2019 |archiveurl=https://web.archive.org/web/20190202131420/https://www.cbc.ca/news/technology/pinocchio-rex-long-snouted-tyrannosaur-discovered-in-asia-1.2634651 |url-status=live}}</ref>。他にも George Olshevsky や Tracy Ford といった研究者が様々なティラノサウルス科恐竜の組み合わせでティラノサウルス亜科の他の下位分類や族を作成したが<ref name="Olshevsky1995">{{Cite journal|last=Olshevsky |first=George |year=1995 |title=The origin and evolution of the tyrannosaurids |journal=Kyoryugaku Saizensen [Dino Frontline] |volume=9–10 |pages=92–119}}</ref><ref name=olshevsky&ford2005>{{cite journal |author1=Olshevsky G. |author2=Ford T.L. | year = 1995 | title = The origin and evolution of the Tyrannosauridae, part 2 [in Japanese] | journal = Dino Frontline | volume = 6 | pages = 75–99 }}</ref>、これらは系統学的に定義されておらず、またこれらを構成する属は他の属や種のシノニムと考えられている<ref name="currieetal2003"/>。 |
|||
|year = 2006 |
|||
|publisher = [[NHK出版|日本放送出版協会]] |
|||
さらなる亜科はより断片的な属にちなんで命名されており、[[アウブリソドン]]亜科と[[デイノドン]]亜科が挙げられる。しかし、アウブリソドンとデイノドンは通例[[疑問名]]と考えられており、そのため彼らや彼らのエポニムの亜科はティラノサウルス科の分類から通常除外される。さらなるティラノサウルス科として、当初はより基盤的なティラノサウルス上科と考えられていたが、タルボサウルスに類似するティラノサウルス亜科の幼体を代表する可能性が高い[[ラプトレックス]]がいる。しかし、本属は幼体の標本しか知られていないため、これも疑問名と考えられている<ref name=fowleretal2011>{{cite journal | last1 = Fowler | first1 = DW | last2 = Woodward | first2 = HN | last3 = Freedman | first3 = EA | last4 = Larson | first4 = PL | last5 = Horner | first5 = JR | year = 2011 | title = Reanalysis of "Raptorex kriegsteini": A Juvenile Tyrannosaurid Dinosaur from Mongolia | journal = PLOS ONE | volume = 6 | issue = 6| page = e21376 | doi = 10.1371/journal.pone.0021376 | bibcode=2011PLoSO...6E1376F | pmid=21738646 | pmc=3126816| doi-access = free }}</ref>。 |
|||
|series = [[NHKブックス]] |
|||
|isbn = 4-14-091061-5 |
|||
=== 系統 === |
|||
|ref = harv |
|||
古脊椎動物学において系統分類学が出現し、ティラノサウルス科は複数の明白な定義を与えられている。最初の定義はポール・セレノによる1998年のもので、[[アレクトロサウルス]]や[[アウブリソドン]]あるいは[[ナノティラヌス]]よりもティラノサウルスに近縁な全てのティラノサウルス上科が本科に含まれた<ref name=sereno1998>{{Cite journal|last=Sereno |first=Paul C. |year=1998 |title=A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria |journal=Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen |volume=210 |pages=41–83|language=de|doi=10.1127/njgpa/210/1998/41 }}</ref>。しかし、ナノティラヌスはよくティラノサウルスの幼体と考えられており、アウブリソドンも[[疑問名]]であるため、系統群の定義には不適である<ref name=holtz2004/>。 |
|||
2001年、トーマス・R・ホルツ・ジュニアはティラノサウルス科の分岐分析を発表した<ref name="holtz2001" />。彼は、鋸歯の無い[[前上顎骨]]歯で特徴づけられる原始的なアウブリソドン亜科と、ティラノサウルス亜科の2亜科が存在すると結論した<ref name="holtz2001" />。アウブリソドン亜科にはアウブリソドンの他に "[[:en:Kirtland Formation|Kirtland]] ''Aublysodon''" と[[アレクトロサウルス]]が含まれる<ref name="holtz2001" />。また、ホルツは[[シャモティラヌス]]がティラノサウルス科の[[共有派生形質]]を複数示すことを発見したが、ティラノサウルス科には置かなかった<ref name="holtz2001" />。 |
|||
同誌で後述された記述にて、彼はティラノサウルス科を「ティラノサウルスとアウブリソドンの[[最も近い共通祖先]]の全ての子孫」として定義することを提唱した<ref name="holtz2001" />。また、彼はポール・セレノのものをはじめ他の研究者が過去に提唱した定義を批判した<ref name="holtz2001" />。ホルツはナノティラヌスがティラノサウルス・レックスの幼体の誤同定であると考えていたため、セレノが提唱した定義ではティラノサウルス科はティラノサウルス属の下位分類になるとしたのである<ref name="holtz2001" />。さらに、セレノの提案した定義ではティラノサウルス亜科もティラノサウルスのみに限定されるとした<ref name="holtz2001">Holtz, TR, Jr. (2001), The phylogeny and taxonomy of the Tyrannosauridae in K Carpenter & D Tanke [eds.], Mesozoic Vertebrate Life. Indiana Univ. Press, pp. 64-83.</ref>。 |
|||
2003年にクリストファー・ブロチュは[[アルバートサウルス]]、[[アレクトロサウルス]]、[[アリオラムス]]、[[ダスプレトサウルス]]、[[ゴルゴサウルス]]、[[タルボサウルス]]およびティラノサウルスを定義に加えた<ref name=Brochu2003b>{{Cite journal|last=Brochu |first=Christopher R. |year=2003 |title=Osteology of ''Tyrannosaurus rex'': insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull |journal=Society of Vertebrate Paleontology Memoirs |volume=7 |pages=1–138 |doi=10.2307/3889334 |jstor=3889334}}</ref>。ホルツは2004年に系統群を再定義し、彼の分析で確固たる位置に置かれなかったアリオラムスとアレクトロサウルスを除いて上記の全てを指示子として用いた。しかし、彼は同じ論文において、[[エオティラヌス]]よりもティラノサウルスに近縁な全ての獣脚類を含むものとして別の定義も提供した<ref name=holtz2004/>。セレノによる2005年の定義では、ティラノサウルス科はアルバートサウルス、ゴルゴサウルス、ティラノサウルスを含む最も包括的でない系統群として定義された<ref name=sereno2005>{{Cite web |url=http://www.taxonsearch.org/dev/file_home.php |title=Stem Archosauria—TaxonSearch |accessdate=14 January 2008 |last=Sereno |first=Paul C. |date=7 November 2005 |archiveurl=https://web.archive.org/web/20070219102659/http://www.taxonsearch.org/dev/file_home.php |archivedate=19 February 2007 }}</ref>。 |
|||
ティラノサウルス科系統の分岐分析では、よくタルボサウルスはティラノサウルスの[[姉妹群]]に置かれ、ダスプレトサウルスはそれらよりも基盤的な位置に置かれる。タルボサウルスとティラノサウルスの近縁性は多数の頭蓋骨の特徴に支持されており、特定の骨の縫合線のパターン、それぞれの目の後ろの後眼窩骨の三日月型をしたクレストの存在、下端が下側に顕著に湾曲する非常に深い上顎骨などがある<ref name=holtz2004/><ref name=carretal2005/>。2003年にフィリップ・J・カリーらが提唱した別の仮説では、鼻骨と涙骨を接続する骨質の先端部が存在しないことに基づき、ダスプレトサウルスはアジア産のタルボサウルスやアリオラムスを含む系統群の基盤的メンバーであるという弱い支持が示された<ref name=currieetal2003/>。 |
|||
関連する研究でも、2属の間で共有される下顎の固定機序が指摘されている<ref name=hurumsabath2003>{{Cite journal |last=Hurum |first=Jørn H. |author2=Sabath, Karol. |year=2003 |title=Giant theropod dinosaurs from Asia and North America: Skulls of ''Tarbosaurus bataar'' and ''Tyrannosaurus rex'' compared |journal=Acta Palaeontologica Polonica |volume=48 |issue=2 |pages=161–190 |url=http://www.app.pan.pl/article/item/app48-161.html?pdf=39 |format=abstract |accessdate=2008-09-10 |archivedate=2012-03-08 |archiveurl=https://web.archive.org/web/20120308104302/http://www.app.pan.pl/article/item/app48-161.html?pdf=39 |url-status=live }}</ref>。別の論文でカリーはアリオラムスがタルボサウルスの幼体である可能性を指摘したが、より卓越したアリオラムスの鼻骨クレストと本数の多い歯から別属であることが示唆されているとも主張した。同様に、カリーはナノティラヌスの歯の本数を用いて同属がティラノサウルスと別属である可能性を示唆した<ref name=currie2003a/>。しかし、[[キアンゾウサウルス]]の発見・記載により、アリオラムスはタルボサウルスと近縁ではなく、新たに記載されたアリオラムス族に属するとされた。さらに、キアンゾウサウルスの存在からは、吻部の長いティラノサウルス科の恐竜がアジアに幅広く分布しており、また異なる獲物を狩ることでより大型かつ頑強なティラノサウルス亜科との競合を回避しつつ環境を共有していたことが明らかになった<ref>{{cite web |url=https://www.sciencedaily.com/releases/2014/05/140507100421.htm |title=Newly found dinosaur is long-nosed cousin of Tyrannosaurus rex |work=ScienceDaily |accessdate=2018-03-09 |archivedate=2018-03-17 |archiveurl=https://web.archive.org/web/20180317102219/https://www.sciencedaily.com/releases/2014/05/140507100421.htm |url-status=live }}</ref>。{{Clear}} |
|||
{| |
|||
|{{Cladogram|title='''Brusatte & Carr, 2016'''<ref>{{sfnRef|Brusatte & Carr|2016}}</ref> |
|||
|clades={{clade| style=font-size:80%;line-height:80% |
|||
|label1='''ティラノサウルス科''' |
|||
|1={{clade |
|||
|label1=[[アルバートサウルス亜科]] |
|||
|1={{clade |
|||
|1=''[[ゴルゴサウルス|Gorgosaurus]]'' |
|||
|2=''[[アルバートサウルス|Albertosaurus]]'' }} |
|||
|label2='''ティラノサウルス亜科''' |
|||
|2={{clade |
|||
|1={{clade |
|||
|label1='''アリオラムス族''' |
|||
|1={{clade |
|||
|1=''[[キアンゾウサウルス|Qianzhousaurus]]'' |
|||
|2={{clade |
|||
|1=''[[アリオラムス|Alioramus remotus]]'' |
|||
|2=''[[アリオラムス|Alioramus altai]]'' }} }} }} |
|||
|2={{clade |
|||
|1={{clade |
|||
|1=''[[ナヌークサウルス|Nanuqsaurus]]'' |
|||
|2=''[[テラトフォネウス|Teratophoneus]]'' }} |
|||
|2={{clade |
|||
|1=''[[リトロナクス|Lythronax]]'' |
|||
|2={{clade |
|||
|1={{clade |
|||
|1=''[[ダスプレトサウルス|Daspletosaurus torosus]]'' |
|||
|2=''[[ダスプレトサウルス|Daspletosaurus horneri]]'' }} |
|||
|2={{clade |
|||
|1=''[[ズケンティラヌス|Zhuchengtyrannus]]'' |
|||
|2={{clade |
|||
|1=''[[タルボサウルス|Tarbosaurus]]'' |
|||
|2=''[[ティラノサウルス|Tyrannosaurus]]'' }} }} }} }} }} }} }} }} }} |
|||
|{{Cladogram|title='''Fiorillo ''et al.'' 2014'''<ref>{{sfnRef|Fiorillo & Tykoski|2014}}</ref> |
|||
|clades={{clade| style=font-size:80%;line-height:80% |
|||
|label1='''ティラノサウルス科''' |
|||
|1={{clade |
|||
|label1=[[アルバートサウルス亜科]] |
|||
|1={{clade |
|||
|1=''[[アルバートサウルス|Albertosaurus]]'' |
|||
|2=''[[ゴルゴサウルス|Gorgosaurus]]'' }} |
|||
|label2='''ティラノサウルス亜科''' |
|||
|2={{clade |
|||
|1={{clade |
|||
|1=''[[ダスプレトサウルス|Daspletosaurus]]'' |
|||
|2=Two Medicine taxon }} |
|||
|2={{clade |
|||
|1=''[[テラトフォネウス|Teratophoneus]]'' |
|||
|2={{clade |
|||
|1=''[[ビスタヒエヴェルソル|Bistahieversor]]'' |
|||
|2={{clade |
|||
|1=''[[リトロナクス|Lythronax]]'' |
|||
|2={{clade |
|||
|1=''[[ナヌークサウルス|Nanuqsaurus]]'' |
|||
|2={{clade |
|||
|1={{clade |
|||
|1=''[[タルボサウルス|Tarbosaurus]]'' |
|||
|2=''[[ズケンティラヌス|Zhuchengtyrannus]]'' }} |
|||
|2=''[[ティラノサウルス|Tyrannosaurus]]'' }} }} }} }} }} }} }} }} }} |
|||
|{{Cladogram|title='''Loewen ''et al.'' 2013'''<ref>{{Cite journal | last1 = Loewen | first1 = M.A. | last2 = Irmis | first2 = R.B. | last3 = Sertich | first3 = J.J.W. | last4 = Currie | first4 = P. J. | author-link4 = フィリップ・J・カリー | last5 = Sampson | first5 = S. D. | year = 2013| title = Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans | editor-last = Evans | editor-first = David C| editor-link = David C. Evans| journal = [[PLoS ONE]] | volume = 8 | issue = 11 | pages = e79420 | doi = 10.1371/journal.pone.0079420 | pmid = 24223179| pmc = 3819173| ref = {{sfnRef|Loewen ''et al.''|2013}}| bibcode = 2013PLoSO...879420L| doi-access = free }}</ref> |
|||
|clades={{clade| style=font-size:80%;line-height:80% |
|||
|label1='''ティラノサウルス科''' |
|||
|1={{clade |
|||
|1={{clade |
|||
|1=''[[ゴルゴサウルス|Gorgosaurus]]'' |
|||
|2=''[[アルバートサウルス|Albertosaurus]]'' }} |
|||
|label2='''ティラノサウルス亜科''' |
|||
|2={{clade |
|||
|1=[[ダイナソーパーク累層|Dinosaur Park]] tyrannosaurid |
|||
|2={{clade |
|||
|1=''[[ダスプレトサウルス|Daspletosaurus]]'' |
|||
|2={{clade |
|||
|1=''[[ダスプレトサウルス|Daspletosaurus horneri]]'' |
|||
|2={{clade |
|||
|1=''[[テラトフォネウス|Teratophoneus]]'' |
|||
|2={{clade |
|||
|1=''[[ビスタヒエヴェルソル|Bistahieversor]]'' |
|||
|2={{clade |
|||
|1=''[[リトロナクス|Lythronax]]'' |
|||
|2={{clade |
|||
|1=''[[ティラノサウルス|Tyrannosaurus]]'' |
|||
|2={{clade |
|||
|1=''[[タルボサウルス|Tarbosaurus]]'' |
|||
|2=''[[ズケンティラヌス|Zhuchengtyrannus]]'' }} }} }} }} }} }} }} }} }} }} |
|||
}} |
}} |
||
|}{{Clear}} |
|||
== 古生物学 == |
|||
=== 成長 === |
|||
{{Annotated image | float=right | caption=Hypothesized growth curves (body mass versus age) of four tyrannosaurids<ref name="ericksonetal2004"/> |
|||
| image=Tyrannosauroid growth curves unlabelled.png | width=370 | image-width=300 | height=250 |
|||
| image-left=30 | image-top=15 |
|||
|annotations= |
|||
{{Annotation|15|5|Body mass (kg)}} |
|||
{{Annotation|5|22|6,000}} |
|||
{{Annotation|5|52|5,000}} |
|||
{{Annotation|5|85|4,000}} |
|||
{{Annotation|5|116|3,000}} |
|||
{{Annotation|5|147|2,000}} |
|||
{{Annotation|5|178|1,000}} |
|||
{{Annotation|85|222|5}} |
|||
{{Annotation|127|222|10}} |
|||
{{Annotation|173|222|15}} |
|||
{{Annotation|216|222|20}} |
|||
{{Annotation|262|222|25}} |
|||
{{Annotation|308|222|30}} |
|||
{{Annotation|158|235|Age (years)}} |
|||
{{Annotation|239|67|{{legend2|black|{{=}} [[ティラノサウルス]]}}}} |
|||
{{Annotation|239|87|{{legend2|#66ff66|{{=}} [[ダスプレトサウルス]]}}}} |
|||
{{Annotation|239|107|{{legend2|blue|{{=}} [[ゴルゴサウルス]]}}}} |
|||
{{Annotation|239|127|{{legend2|red|{{=}} [[アルバートサウルス]]}}}} |
|||
}} |
|||
古生物学者グレゴリー・エリクソンらはティラノサウルス科の成長と生活史を研究した。ボーンヒストロジーの分析からは標本が死亡した際の年齢を決定でき、様々な個体の年齢を彼らの大きさに対してグラフ上にプロットすれば成長率を調査できる。エリクソンは、ティラノサウルスが長い幼体期間を過ごした後、一生涯の途中の約4年間に驚異的な成長期を迎えたことを明らかにした。性成熟とともに急成長期が終わると、成体では成長がかなり鈍化する。ティラノサウルス類の成長曲線はS字型であり、14歳前後での個体の成長速度が最大となる<ref name=ericksonetal2004/>。 |
|||
[[File:Tyrannosaurus Rex Jane.jpg|alt=|left|thumb|[[ジェーン (ティラノサウルス)|ジェーン]]の骨格のキャスト]] |
|||
既知のティラノサウルス・レックスの個体で最小のものは LACM 28471であるが、これは体重僅か29.9キログラムの2歳の個体である。この一方、最大級の個体の1つとされているものが [[フィールド自然史博物館|FMNH]] PR2081([[スー (ティラノサウルス)|スー]])であり、体重は約5654キログラム、年齢は28歳と推定されており、本種の上限に近かった可能性がある<ref name=ericksonetal2004>{{Cite journal|author=Erickson, Gregory M. |author2=Makovicky, Peter J. |author3=フィリップ・J・カリー |author4=Norell, Mark A. |author5=Yerby, Scott A. |author6=Brochu, Christopher A. |year=2004 |title=Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs |journal=Nature |volume=430 |issue=7001 |pages=772–775 |doi=10.1038/nature02699 |pmid=15306807 |first1=GM|quote=Sampled longevities for T. rex ranged from 2 to 28 years|bibcode=2004Natur.430..772E |s2cid=4404887 }}</ref>。ティラノサウルス・レックスの幼体は約14歳になるまで体重1800キログラム未満であり、約14歳で劇的に体サイズを増大する。この急速な成長期の間に、若いティラノサウルス・レックスは4年間で年間平均600キログラムの体重増加を経験する。この成長速度は16歳から低下し、18歳で再びプラトーに達して劇的に成長が減速する<ref name="hornerpadian2004">{{Cite journal|author1=Horner, J.R. |author2=Padian, K. |name-list-style=amp |year=2004|title=Age and growth dynamics of Tyrannosaurus rex|journal=Proceedings of the Royal Society of London B |pmid=15347508 |volume=271 |issue=1551 |pmc=1691809 |pages=1875–1880 |doi=10.1098/rspb.2004.2829}}</ref>。例えば、28歳のスーと22歳のカナダの標本([[ロイヤル・ティレル古生物学博物館|RTMP]] 81.12.1)の間では僅か600キログラムの差異しかない<ref name="ericksonetal2004"/>。この急激な成長速度変化は肉体的な成熟を示唆する可能性がある。この仮説は[[モンタナ州]]産の18歳のティラノサウルス・レックスの標本(MOR 1125、[[Bレックス]])の大腿骨から骨髄組織が発見されたことで支持されている<ref name=LW08>{{Cite journal|last=Lee |first=Andrew H. |author2=Werning, Sarah |year=2008 |title=Sexual maturity in growing dinosaurs does not fit reptilian growth models |journal=Proceedings of the National Academy of Sciences |volume=105 |issue=2 |pages=582–587 |doi=10.1073/pnas.0708903105 |pmid=18195356 |pmc=2206579|bibcode=2008PNAS..105..582L |doi-access=free }}</ref>。骨髄組織は[[排卵]]期の雌の鳥類でのみ確認されており、このことからBレックスは繁殖期にあったことが示唆される<ref name="schweitzeretal2005">{{Cite journal|author=Schweitzer, M.H., Wittmeyer, J.L., & Horner, J.R. |year=2005 |title=Gender-specific reproductive tissue in ratites and ''Tyrannosaurus rex'' |journal=Science|volume=308|pages=1456–1460 |doi=10.1126/science.1112158 |pmid=15933198 |issue=5727 |bibcode=2005Sci...308.1456S |s2cid=30264554 |url=https://semanticscholar.org/paper/33a27d3e689b35f6cce1b37ac37bc7e5aec31074 |accessdate=2019-12-02 |archivedate=2021-11-10 |archiveurl=https://web.archive.org/web/20211110091111/https://www.semanticscholar.org/paper/Gender-Specific-Reproductive-Tissue-in-Ratites-and-Schweitzer-Wittmeyer/33a27d3e689b35f6cce1b37ac37bc7e5aec31074 }}</ref>。 |
|||
他のティラノサウルスも酷似した成長曲線を示すが、成体のサイズがティラノサウルスと比較して小さいため、成長速度もそれに対応する<ref name="ericksonetal2006">{{Cite journal|author=Erickson, G.M., Currie, P.J., Inouye, B.D., & Winn, A.A. |year=2006 |title=Tyrannosaur life tables: an example of nonavian dinosaur population biology |journal=Science |volume=313 |pages=213–217 |doi=10.1126/science.1125721 |pmid=16840697 |issue=5784 |bibcode=2006Sci...313..213E |s2cid=34191607 |url=https://semanticscholar.org/paper/675afac6b97ad6f810f5daf7d1633434d548f0fc|accessdate=2019-12-02 |archivedate=2021-11-10 |archiveurl=https://web.archive.org/web/20211110091104/https://www.semanticscholar.org/paper/Tyrannosaur-Life-Tables%3A-An-Example-of-Nonavian-Erickson-Currie/675afac6b97ad6f810f5daf7d1633434d548f0fc}}</ref>。アルバートサウルス亜科と比較してダスプレトサウルスは急速成長期における成長速度が高いが、これは成体の体重がより重いためである。成体体重1800キログラムという推定に基づくと、ダスプレトサウルスの最大成長速度は年間180キログラムである。他の研究者はダスプレトサウルスの体重をより重く見積もっているため成長速度の規模は左右されうるが、全体のパターンに変化はないと考えられる<ref name=ericksonetal2004/>。最も若い既知のアルバートサウルスはドライアイランドのボーンベッドで発見された2歳の個体で、体重は約50キログラム、と推定され、全長は2メートルを僅かに上回る。同じ産地からは既知で最大かつ最高齢の個体が発見されており、全長は10メートル、年齢は28歳である。最速成長速度は12 - 16歳で迎えたと推定され、体重1300キログラムの成体に基づき、その速度は年間122キログラムに達したと見積られる。これはティラノサウルス・レックスの成長速度の五分の一である。ゴルゴサウルスでは、計算された最大成長速度は急成長段階において約110キログラムであり、アルバートサウルスのものに匹敵する<ref name=ericksonetal2004/>。 |
|||
いまだ未確認の属のティラノサウルス類の[[胚]]が発見されており、ティラノサウルス科は卵の中での発生段階で特徴的な骨格構造を発達させることが示唆される。さらに、1983年にモンタナ州のツーメディスン層で発見された下顎の1.1インチ(2.8cm)の歯と、2018年にホースシューキャニオン層で発見され2020年に記載された足の鉤爪の標本の大きさから、ティラノサウルス科の新生児はマウスやそれに近い大きさの齧歯類の全身と同等規模の頭蓋骨を伴って生まれ、全身はほぼ小型犬程度の大きさだった可能性があると考えられる。下顎の標本は全長約0.76メートルの個体、鉤爪は全長約0.91メートルの個体のものであると考えられている。いずれの標本も関連する卵殻は未発見であるが、これらのティラノサウルス科の新生児が発見された場所はこの動物が共存・捕食した他の種と同じ巣を利用していたことが示唆される<ref>{{Cite web|website=Science |accessdate=November 23, 2020 |archivedate=December 5, 2020 |url=https://www.nationalgeographic.com/science/2020/10/first-tyrannosaur-embryo-fossils-revealed/ |title=First tyrannosaur embryo fossils revealed|date=October 19, 2020 |archiveurl=https://web.archive.org/web/20201205015329/https://www.nationalgeographic.com/science/2020/10/first-tyrannosaur-embryo-fossils-revealed/ }}</ref>。また、卵殻がこれらの標本に無いことから、ティラノサウルス科は[[ムササウルス]]属や[[プロトケラトプス]]属がそう考えられているように、軟殻の卵を産んだ可能性が指摘されている<ref>{{Cite web|url=https://www.livescience.com/baby-embryonic-tyrannosaur-fossils.html |title=1st of their kind baby tyrannosaur fossils unearthed |first=Laura Geggel-Associate Editor 15 |last=October 2020 |website=livescience.com |date=15 October 2020 |accessdate=17 October 2020 |archivedate=17 October 2020 |archiveurl=https://web.archive.org/web/20201017201213/https://www.livescience.com/baby-embryonic-tyrannosaur-fossils.html}}</ref>。 |
|||
ワピチ層の足跡化石からは、ティラノサウルス科は爪先が厚くなって足が幅広になり、体重を支持するように成長したことが示唆される。幅広の足は、成体のティラノサウルス科が子孫よりも緩慢に動いていたことを示唆する<ref>{{Cite web|url=http://theconversation.com/fat-footed-tyrannosaur-parents-couldnt-keep-up-with-their-skinnier-offspring-fossil-footprints-reveal-154188 |title=Fat-footed tyrannosaur parents couldn't keep up with their skinnier offspring, fossil footprints reveal|first1=Nathan James |last1=Enriquez |first2=Nicolas |last2=Campione |first3=Phil |last3=Bell |website=The Conversation |accessdate=2021-04-28 |archivedate=2021-04-26 |archiveurl=https://web.archive.org/web/20210426230718/https://theconversation.com/fat-footed-tyrannosaur-parents-couldnt-keep-up-with-their-skinnier-offspring-fossil-footprints-reveal-154188 }}</ref><ref>{{Cite journal|title=Exploring possible ontogenetic trajectories in tyrannosaurids using tracks from the Wapiti Formation (upper Campanian) of Alberta, Canada |first1=Nathan J. |last1=Enriquez |first2=Nicolás E. |last2=Campione |first3=Tom |last3=Brougham |first4=Federico |last4=Fanti |first5=Matt A. |last5=White |first6=Robin L. |last6=Sissons |first7=Corwin |last7=Sullivan |first8=Matthew J. |last8=Vavrek |first9=Phil R. |last9=Bell |date=April 21, 2021 |journal=Journal of Vertebrate Paleontology |volume=40 |issue=6 |pages=e1878201 |doi=10.1080/02724634.2021.1878201 |doi-access=free}}</ref>。 |
|||
=== 生活史 === |
|||
[[Image:Gorgosaurus death pose.jpg|thumb|ゴルゴサウルス・リブラトゥスのほぼ完全な骨格。[[ロイヤル・ティレル古生物学博物館]]]] |
|||
急成長段階の終わりからは、アルバートサウルスは性成熟期に入ったことが示唆される。ただし、その後も一生涯を通じて緩慢に成長を続けたことも示唆される<ref name=ericksonetal2004/><ref name=ericksonetal2006/>。まだ活発に成長している時期に性的に成熟することは小型<ref name=ericksonetal2007>{{Cite journal|last1=Erickson |first1=Gregory M. |last2= Curry Rogers|first2=Kristi |last3=Varricchio |first3=David J. |last4=Norell |first4=Mark |last5=Xu |first5=Xing |year=2007 |title=Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition |journal=Biology Letters |volume=3 |pages=558–561 |doi=10.1098/rsbl.2007.0254 |pmid=17638674 |issue=5 |pmc=2396186}}</ref>あるいは大型<ref name=lee_werning2008>{{Cite journal|last=Lee |first=Andrew H. |author2=Werning, Sarah |year=2008 |title=Sexual maturity in growing dinosaurs does not fit reptilian growth models |journal=PNAS |volume=105 |pages=582–587 |doi=10.1073/pnas.0708903105 |pmid=18195356 |issue=2 |pmc=2206579|bibcode=2008PNAS..105..582L |doi-access=free }}</ref>恐竜だけでなく、ヒトや[[ゾウ]]といった大型哺乳類にも共通する形質のようである<ref name=lee_werning2008/>。相対的に性成熟の早いパターンは、成長が完了した後まで性成熟を遅延させる鳥類のパターンとは顕著に異なる<ref name=lee_werning2008/><ref name=ricklefs2007>{{Cite journal|last=Ricklefs |first=Robert E. |year=2007 |title=Tyrannosaur ageing |journal=Biology Letters |volume=3 |issue=2 |pages=214–217 |doi=10.1098/rsbl.2006.0597 |pmid=17284406 |pmc=2375931}}</ref>。 |
|||
各年齢層の標本数を集計することにより、エリクソンらはティラノサウルス科の個体群における生活史について結論を導いた。その結果、化石の記録では幼体は稀である一方、急成長期にある亜成体や成体は遥かに多いことが判明した。半数を上回るティラノサウルス・レックスの標本は性成熟後6年以内に死亡しているようであり、これは他のティラノサウルス類や、現代の大型かつ長寿の鳥類や哺乳類にも見られるパターンである。これらの種は新生児の時期の死亡率が高く、次いで幼体の時期の死亡率が比較的低いことが特徴である。性成熟後には死亡率が再び上昇するが、これは生殖のストレスが一因である。これは保存状態や採集のバイアスに起因する可能性もあるが、エリクソンは、ゾウなど現代の大型哺乳類にも見られる一定以上の大きさの幼体の死亡率が低いことが原因ではないかと推論した。ティラノサウルスは2歳までに同時代の全ての捕食者を上回る大きさになるため、捕食者の不在に起因して死亡率が低下した可能性がある。古生物学者が十分な数のダスプレトサウルスの化石を発見していないため同様の分析は不可能であるが、エリクソンは同様の一般的な傾向があるようだと指摘した<ref name=ericksonetal2006/>。 |
|||
ティラノサウルス科はその生涯の半分を幼体として過ごした後、わずか数年でほぼ最大サイズまで急成長を遂げる<ref name=ericksonetal2004/>。このことと、巨大な成体のティラノサウルス科と他の小型獣脚類の中間的な大きさの捕食者が完全に欠落していることから、これらの生態的地位は幼体のティラノサウルス科が埋めていたことが示唆される。これは孵化した後に樹上生活を送り、緩慢に成熟して大型脊椎動物を引き倒すことの出来る頑丈な[[頂点捕食者]]に至る、現生の[[コモドオオトカゲ]]にも見られる<ref name=holtz2004/>。例えば、アルバートサウルスは群集で発見されているが、その群れには異なる年齢の個体が入り混じっている<ref name=tankecurrie1998/><ref name=currieetal2005/>。 |
|||
=== 移動 === |
|||
移動能力が最もよく研究されているのはティラノサウルスについてであり、これに関して2つの話題がある。1つはどれほど旋回が得意であったか、もう1つは直線運動の最高速度はどれほどであったか、である。ティラノサウルスの旋回は緩慢であった可能性があり、45度回転するのに1~2秒を要したと考えられる。45度とは、直立していてかつ尾のない人間が1秒で回転可能な量である<ref>{{cite web |url=http://www.cosmosmagazine.com/node/1381 |title=Tyrannosaurus had poor turning circle |archivedate=May 7, 2012 |accessdate=2007-09-25 |archiveurl=https://web.archive.org/web/20120507053958/http://www.cosmosmagazine.com/node/1381}}</ref>。旋回を苦手とする原因は回転慣性にある。これは、ティラノサウルスの質量の多くは、人間が重い材木を運ぶように、重心からある程度離れたところに存在したためである<ref name="CarrierWalterLee2000TurningPerformance">{{Cite journal | title=Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia | journal=Journal of Experimental Biology | archivedate=6 January 2008 | issue=Pt 22 | pages=3917–3926 | url=http://jeb.biologists.org/cgi/content/full/204/22/3917 | author=Carrier, D.R., Walter, R.M., and Lee, D.V. | pmid=11807109 | date=15 November 2001 | volume=204 | doi=10.1242/jeb.204.22.3917 | accessdate=12 October 2008 | archiveurl=https://web.archive.org/web/20080106180207/http://jeb.biologists.org/cgi/content/full/204/22/3917 }}</ref>。 |
|||
研究者による最大速度の推定は非常に値の幅が広く、主に約11メートル毎秒と推定されているが、5 - 11メートル毎秒や20メートル毎秒とする推定値もある。この原因は、歩行した痕跡は数多く発見されている一方で走行した痕跡は発見されておらず、彼らが走行しなかったことを示す可能性があるからである<ref name="Hutchinson, J.R. 2004 441–461">{{Cite journal |author=Hutchinson, J.R. |title=Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa |journal=Journal of Morphology |volume=262 |issue=1 |pages=441–461 |year=2004 |url=http://www.rvc.ac.uk/AboutUs/Staff/jhutchinson/documents/JRH13.pdf |doi=10.1002/jmor.10240 |pmid=15352202 |s2cid=15677774 |archiveurl=https://web.archive.org/web/20081031093050/http://www.rvc.ac.uk/AboutUs/Staff/jhutchinson/documents/JRH13.pdf |archivedate=2008-10-31 }}</ref>。 |
|||
[[ファイル:T Rex Leg Bones.png|サムネイル|左|ティラノサウルス・レックスの後肢]] |
|||
1993年、ジャック・ホーナーとドン・セッレムはティラノサウルスが緩慢でありおそらく走行できなかった(脚が空中に浮かぶフェーズが存在しない)と主張した<ref name="hornerlessem1993">{{Cite book |first=John R. |last=Horner |author2=Don Lessem |title=The complete T. rex |publisher=Simon & Schuster |location=New York City |year=1993 |pages=[https://archive.org/details/completetrexhows00horn/page/120 120] |isbn=978-0-671-74185-3 |url=https://archive.org/details/completetrexhows00horn/page/120 }}</ref>。しかし、Holtz (1998) はティラノサウルス科およびその近縁な動物は最速の大型獣脚類だったと結論した<ref name="Holtz1998TaxonomyCoelurosauria">{{Cite journal |first=Thomas R. |last=Holtz |title=Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda) |journal=Journal of Paleontology |date=1 May 1996 |volume=70 |issue=3 |pages=536–538 |url=http://jpaleontol.geoscienceworld.org/cgi/content/abstract/70/3/536 |accessdate=3 October 2008 |doi=10.1017/S0022336000038506 |s2cid=87599102 |archivedate=26 October 2008 |archiveurl=https://web.archive.org/web/20081026053239/http://jpaleontol.geoscienceworld.org/cgi/content/abstract/70/3/536 }}</ref>。Christiansen (1998) はティラノサウルスの脚の骨がトップスピードが限定されていてかつ実際に走行できないゾウのものよりも遥かに強靭であるわけではないと推定し、ティラノサウルスの最高速度を約11メートル毎秒と提唱した。これはヒトのスプリンターの速度と同程度である<ref name="Christiansen1998Strength">{{Cite journal |author=Christiansen, P. |title=Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential |journal=Gaia |year=1998 |volume=15 |pages=241–255 |url=http://www.mnhn.ul.pt/geologia/gaia/19.pdf |issn=0871-5424 |accessdate=2008-10-12 |archivedate=2008-10-31 |archiveurl=https://web.archive.org/web/20081031093050/http://www.mnhn.ul.pt/geologia/gaia/19.pdf }}</ref>。Farlow ほか (1995) は6 - 8トンに達するティラノサウルスが高速運動中に転倒すれば、地面に叩き付けられた際に小さな前肢では衝撃を減衰させられず重症あるいは致命傷を負ったと主張した<ref name=farlowetal1995>{{Cite journal |doi=10.1080/02724634.1995.10011257 |last=Farlow |first=James O. |author-link=James Farlow |author2=Smith, Matthew B. |author3=Robinson, John M. |year=1995 |title=Body mass, bone "strength indicator", and cursorial potential of ''Tyrannosaurus rex'' |journal=Journal of Vertebrate Paleontology |volume=15 |issue=4 |pages=713–725 |url=http://www.vertpaleo.org/publications/jvp/15-713-725.cfm |archiveurl=https://web.archive.org/web/20081023063102/http://www.vertpaleo.org/publications/jvp/15-713-725.cfm |archivedate=2008-10-23 }}</ref><ref>[https://www.newscientist.com/article/mg14819982.600-the-bigger-they-come-the-harder-they-fall.html "The bigger they come, the harder they fall"] {{Webarchive|url=https://web.archive.org/web/20150531201705/http://www.newscientist.com/article/mg14819982.600-the-bigger-they-come-the-harder-they-fall.html |date=2015-05-31 }} New Scientist, 7 October 1995, p. 18.</ref>。しかし、[[動物園]]のような安全な環境に置かれていたとしても、[[キリン]]は骨折の危険がありながらも50キロメートル毎時でギャロップを行うことが知られている<ref>{{Cite web |url=http://www.wildlifesafari.info/giraffe.html |publisher=WildlifeSafari.info |title=Giraffe |accessdate=29 April 2006 |archivedate=12 June 2010 |archiveurl=https://web.archive.org/web/20100612080401/http://www.wildlifesafari.info/giraffe.html |url-status=live }}</ref><ref>{{Cite web|url=http://www.zoo.org/zoo_info/history/chapters/history4.html |title=The History of Woodland Park Zoo — Chapter 4 |accessdate=29 April 2006 |archiveurl=https://web.archive.org/web/20070602141617/http://www.zoo.org/zoo_info/history/chapters/history4.html |archivedate=June 2, 2007 }}</ref>。このため、ティラノサウルスも必要に応じて高速で動き、そのような危険を受け入れていた可能性もある。このシナリオは[[アロサウルス]]においても研究されている<ref name="Alexander2006DinoBioMechanics">{{Cite journal|author=Alexander, R.M. |title=Dinosaur biomechanics |journal=Proc Biol Sci |date=August 7, 2006 |volume=273 |issue=1596 |pages=1849–1855 |doi=10.1098/rspb.2006.3532 |pmid=16822743 |pmc=1634776}}</ref><ref name="Hanna2002MultipleInjuriesBigAl">{{Cite journal|last=Hanna |first=Rebecca R. |year=2002 |title=Multiple injury and infection in a sub-adult theropod dinosaur (''Allosaurus fragilis'') with comparisons to allosaur pathology in the Cleveland-Lloyd Dinosaur Quarry collection |journal=Journal of Vertebrate Paleontology |volume=22 |issue=1 |pages=76–90 |doi=10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2|s2cid=85654858 |issn=0272-4634}}</ref>。さらに後の研究では、ティラノサウルスの走行速度は17キロメートル毎時から40キロメートル毎時、すなわち歩行や緩慢な走行から中速の走行までで、それ以上に絞り込まれてはいない<ref name="Hutchinson, J.R. 2004 441–461"/><ref name="HutchinsonGarcia2002TrexSlow">{{Cite journal|author8=Hutchinson, J. R. and Garcia, M. |date=28 February 2002 |title=''Tyrannosaurus'' was not a fast runner |journal=Nature |volume=415 |pages=1018–1021 |doi=10.1038/4151018a |author=Hutchinson, John R. |pmid=11875567 |last2=Garcia |first2=M |issue=6875 |bibcode=2002Natur.415.1018H|s2cid=4389633 }}</ref><ref name="dinocards7">Hajdul, R. (1997). Tendons. ''Dinosaur Cards''. Orbis Publishing Ltd. D36044311.</ref>。化石から直接得られたデータを用い2007年にコンピュータモデル研究によって推定された走行速度は8メートル毎秒であった<ref name="SellersManning2007ProcRSocB">{{Cite journal |title=Estimating dinosaur maximum running speeds using evolutionary robotics |journal=Proc. R. Soc. B |doi=10.1098/rspb.2007.0846 |date=July 2007 |url=http://dinonews.net/rubriq/articles.php5?action=open&ref=2007_sellers_speed |author1=Sellers, W.I. |author2=Manning, P.L. |name-list-style=amp |volume=274 |pmid=17711833 |issue=1626 |pmc=2279215 |pages=2711–6 |accessdate=2008-10-12 |archivedate=2011-02-07 |archiveurl=https://web.archive.org/web/20110207142321/http://dinonews.net/rubriq/articles.php5?action=open&ref=2007_sellers_speed }}</ref><ref>[http://news.bbc.co.uk/2/hi/science/nature/6956867.stm Liz Seward: "T. rex 'would outrun footballer'"] {{Webarchive|url=https://web.archive.org/web/20120107031152/http://news.bbc.co.uk/2/hi/science/nature/6956867.stm |date=2012-01-07 }}. ''BBC News'' website, Tuesday, 21 August 2007. The article quotes Dr Bill Sellers, University of Manchester, co-author of a paper published in ''Proceedings of the Royal Society B''. Retrieved 22 August 2007.</ref>。 |
|||
Eric Snively らが2019年に発表した研究では、タルボサウルスやティラノサウルスといったティラノサウルス科は同サイズの[[アロサウルス上科]]よりも機動性が高いことが示唆された。これは、大型の後肢の筋肉に結び付く体重と比較して回転慣性が小さいためである。結果として、ティラノサウルス科は相対的に素早い旋回が可能で、獲物に近づく際あるいは他に旋回する際にはそういった動作を見せた可能性が高く、または旋回中に片脚を固定しながらもう片方を持ち上げる動作をした可能性があるとの仮説が立てられた。この研究結果は、ティラノサウルス類の進化に敏捷性が寄与した過程を明らかにする可能性がある<ref>{{Cite journal|title=Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods |first1=Eric |last1=Snively |first2=Haley |last2=O’Brien |first3=Donald M. |last3=Henderson |first4=Heinrich |last4=Mallison |first5=Lara A. |last5=Surring |first6=Michael E. |last6=Burns |first7=Thomas R. Holtz |last7=Jr |first8=Anthony P. |last8=Russell |first9=Lawrence M. |last9=Witmer |first10=Philip J. |last10=Currie |first11=Scott A. |last11=Hartman |first12=John R. |last12=Cotton |date=February 21, 2019 |journal=PeerJ |volume=7 |pages=e6432 |doi=10.7717/peerj.6432 |pmid=30809441 |pmc=6387760 |doi-access=free}}</ref>。 |
|||
加えて、2020年の研究ではティラノサウルス科は例外的に効率的な歩行者であったことが示唆された。Dececchi ''et al.'' によると、ティラノサウルス科を含む70種を超える獣脚類恐竜の後肢・体重・足取りが比較された。調査チームは様々な方法を適用してそれぞれの恐竜の最高速度、および歩行時のように緩やかな速度での運動時に消費したエネルギー量を推定した。[[ドロマエオサウルス科]]などより小型から中型の種の間では、より長い後肢はより速い走行への適応のように見え、他の研究者による以前の結果と整合する。しかし、体重1000キログラムを超える獣脚類では、最高走行速度は体サイズによる制限を受けており、そのためより長い後肢は速度ではなく低エネルギーの歩行に相関することが判明した。さらに研究結果からは、より小型の獣脚類は進化した長い脚を狩りと捕食者からの逃走の両方に役立てていた一方、大型の捕食性獣脚類は頂点捕食者としての役割ゆえに捕食圧から解放されており、進化した長い脚をエネルギーコストの減少と採餌の効率に役立てていたことが示唆された。研究でより基盤的な獣脚類のグループと比較されたティラノサウルス科は、狩りと死肉漁りの間のエネルギー支出が減少したため、採餌効率の顕著な上昇を示した。これは結果としてティラノサウルス類が彼ら自身を維持するために必要な狩りの襲撃と食餌の量を低減することに帰結した可能性が高い。加えて彼らの研究では、ティラノサウルス類が他の大型獣脚類よりもより俊敏であったことを示す研究との繋がりの中で、長距離に亘って獲物へ忍び寄って接近した後に突発的な急スピードで以て殺害することにティラノサウルス類が長けていることが示唆されている。結果としてティラノサウルス科と現生の[[オオカミ]]の間に類似性が指摘されており、これはアルバートサウルスなどの少なくとも一部のティラノサウルス科が集団で狩りを行っていたことによって裏付けられている<ref>{{Cite journal|doi = 10.1371/journal.pone.0223698|title = The fast and the frugal: Divergent locomotory strategies drive limb lengthening in theropod dinosaurs|year = 2020|last1 = Dececchi|first1 = T. Alexander|last2 = Mloszewska|first2 = Aleksandra M.|last3 = Holtz|first3 = Thomas R.|last4 = Habib|first4 = Michael B.|last5 = Larsson|first5 = Hans C. E.|journal = PLOS ONE|volume = 15|issue = 5|pages = e0223698|pmid = 32401793|pmc = 7220109|bibcode = 2020PLoSO..1523698D|doi-access = free}}</ref><ref>{{Cite web|url=https://www.eurekalert.org/pub_releases/2020-05/p-trw051320.php |title=T. rex was a champion walker, super-efficient at lower speeds |website=EurekAlert! |accessdate=2020-06-27 |archivedate=2020-06-27 |archiveurl=https://web.archive.org/web/20200627052208/https://www.eurekalert.org/pub_releases/2020-05/p-trw051320.php|url-status=live}}</ref>。 |
|||
=== 外皮 === |
|||
[[File:Gorgosaurus.png|thumb|系統ブラケッティングによるゴルゴサウルスの復元]] |
|||
古生物学コミュニティで続いている議論として、ティラノサウルス科の外皮の被覆の範囲と特性に関するものがある。[[前期白亜紀]]にあたる{{仮リンク|義県累層|en|Yixian Formation}}や中国の[[遼寧省]]に分布するその他の層からは、無数のコエルロサウルス類の骨格と共に長いタンパク質繊維が保存され、産出している<ref name=zhouetal2003>{{Cite journal|last1=Zhou Zhonghe |author2=Barrett, Paul M. |author3= Hilton, Jason. |year=2003 |title=An exceptionally preserved Lower Cretaceous ecosystem |journal=Nature |volume=421 |issue=6925 |pages=807–814 |doi=10.1038/nature01420 |pmid=12594504 |first1=Z|bibcode=2003Natur.421..807Z |s2cid=4412725 }}</ref>。他の仮説も提唱されているものの<ref name=linghamsoliaretal2007>{{Cite journal|last1=Lingham-Soliar |first1=Theagarten |last2=Feduccia |first2=Alan |last3=Wang |first3=Xiaolin |year=2007 |title=A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres |journal=Proceedings of the Royal Society of London. Series B, Biological Sciences |pmid=17521978 |volume=274 |issue=1620 |pmc=2270928 |pages=1823–1829 |doi=10.1098/rspb.2007.0352}}</ref>、通常これらの繊維は[[鳥類]]および非鳥類型獣脚類に見られる枝分かれした[[羽毛]]とのホモログであるプロトフェザーとして解釈されている<ref name=chenetal1998>{{Cite journal|last1=Chen Peiji |author2=Dong Zhiming |author3= Zhen Shuonan. |year=1998 |title=An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China |journal=Nature |volume=391 |issue=6663 |pages=147–152 |doi=10.1038/34356 |first1=Pei-ji |bibcode=1998Natur.391..147C|s2cid=4430927 |author2-link=Dong Zhiming }}</ref><ref name=xuetal2003>{{Cite journal|author=Xu Xing |author-link=徐星 |author2=Zhou Zhonghe |last3=Prum |first3=Richard A. |year=2003 |title=Branched integumental structures in Sinornithosaurus and the origin of feathers |journal=Nature |volume=410 |issue=6825 |pages=200–204 |doi=10.1038/35065589 |pmid=11242078 |first1=X |bibcode=2001Natur.410..200X|s2cid=4426803 }}</ref>。2004年に記載された[[ディロング]]の骨格はプロトフェザーを含んでおり、これはティラノサウルス上科において確認された最初の例である。現生鳥類の{{仮リンク|ダウンフェザー|en|Down feather}}と同様にディロングに見られたプロトフェザーは枝分かれしていたものの、大羽{{enlink|Pennaceous feather|a=on}}にはなっておらず、体温の保温に用いられていた可能性がある<ref name=xuetal2004/>。2012年には全長9メートルに達する[[ユウティラヌス]]が記載され、本属の発見からは大型獣脚類の成体も羽毛が生えていた可能性が示唆された<ref>{{cite web |url=http://www.xinglida.net/pdf/Xu_et_al_2012_Yutyrannus.pdf |title=Archived copy |accessdate=2012-04-04 |url-status=dead |archiveurl=https://web.archive.org/web/20120417134949/http://www.xinglida.net/pdf/Xu_et_al_2012_Yutyrannus.pdf |archivedate=2012-04-17 }}</ref>。 |
|||
[[File:Royal Tyrrell Museum 2014.6.JPG|thumb|羽毛の生えたアルバートサウルスの彫像]] |
|||
{{仮リンク|系統ブラケッティング|en|Phylogenetic bracketing}}の原則に基づき、ティラノサウルス科はそのような羽毛を持った可能性があると予想された。しかし2017年の研究では、アルバータ州・モンタナ州および[[モンゴル国]]で収集された5属(ティラノサウルス、アルバートサウルス、ゴルゴサウルス、ダスプレトサウルス、タルボサウルス)のティラノサウルス科の皮膚の印象化石が記載された。皮膚の印象化石は小型であるものの、これらは頭部よりも後側に幅広く分布しており、腹部・胸部・腸骨・骨盤・尾部・頸部に位置していた。これらは重複していない細かく小石様の密な鱗を示しており、羽毛の痕跡は無かった。基本的なテクスチャーは直径約1 - 2mmの小さな "basement scales" で構成され、その間に約7mmの "feature scales" の印象が散見される<ref name="rsbl.royalsocietypublishing.org">{{cite journal | last1 = Bell | first1 = P. R. | last2 = Campione | first2 = N. E. | last3 = Persons | first3 = W. S. | last4 = Currie | first4 = P. J. | last5 = Larson | first5 = P. L. | last6 = Tanke | first6 = D. H. | last7 = Bakker | first7 = R. T. | year = 2017 | title = Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution | journal = Biology Letters | volume = 13 | issue = 6| page = 20170092 | doi = 10.1098/rsbl.2017.0092 | pmid = 28592520 | pmc = 5493735 | doi-access = free }}</ref>。ティラノサウルス類の足跡にもさらなる鱗が見られる<ref>Currie, P.J., Badamgarav, D., Koppelhus, E.B. 2003. The First Late Cretaceous Footprints from the Nemegt Locality in the Gobi of Mongolia. Ichnos. Vol.10:1-12.Currie, P.J., Badamgarav, D., Koppelhus, E.B. 2003.</ref>。ティラノサウルス類の顔面の外皮は歯骨と上顎骨に鱗があり、表皮は角質化し、下位領域には鎧のような皮膚があることが研究で判明している<ref>Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. ''Scientific Reports'', 7.</ref><ref>{{Cite web |last1=Witton |first1=Mark |last2=Hone |first2=David |date=September 2018 |title=Tyrannosaurid theropods: did they ever smile like crocodiles? |url=https://iris.unito.it/bitstream/2318/1703198/1/SVPCA2018AbstractBooklet.pdf |website=The Annual Symposium of Vertebrate Palaeontology and Comparative Anatomy |accessdate=2022-10-11}}</ref>。 |
|||
Bell ''et al.'' はティラノサウルス上科の外皮の分布について既知の内容に基づいて祖先形質復元を行った。ティラノサウルス上科の起源に羽毛があることの信頼性は89%であったにも拘わらず、ティラノサウルス科が鱗を持つ信頼性は97%で真と判断された。研究チームはこのデータが「ティラノサウルスが完全に扁平上皮で覆われていたことの説得力ある証拠をもたらす」とした。ただし、皮膚の印象が未発見である背中側にはまだ羽毛が存在していた可能性があると彼らは認めている。Bell ''et al.'' は、裏付ける根拠を必要としつつも、ティラノサウルス類の鱗の印象が羽毛から二次的に派生したものである可能性があるという仮説を立てている<ref name="rsbl.royalsocietypublishing.org"/>。しかし、ティラノサウルス科におけるタフォノミーバイアスに起因するという主張もなされている<ref>{{Cite journal |last1=Saitta |first1=Evan T. |last2=Fletcher |first2=Ian |last3=Martin |first3=Peter |last4=Pittman |first4=Michael |last5=Kaye |first5=Thomas G. |last6=True |first6=Lawrence D. |last7=Norell |first7=Mark A. |last8=Abbott |first8=Geoffrey D. |last9=Summons |first9=Roger E. |last10=Penkman |first10=Kirsty |last11=Vinther |first11=Jakob |date=2018-11-01 |title=Preservation of feather fibers from the Late Cretaceous dinosaur Shuvuuia deserti raises concern about immunohistochemical analyses on fossils |url=https://dspace.mit.edu/bitstream/handle/1721.1/125319/Saitta_et_al_feather_fibres_OG_AAM.pdf?sequence=2&isAllowed=y |journal=Organic Geochemistry |language=en |volume=125 |pages=142–151 |doi=10.1016/j.orggeochem.2018.09.008 |s2cid=105753275 |issn=0146-6380}}</ref>。 |
|||
そのような外皮が何故発生したかはまだ決定されていない。羽毛が喪失した前例は[[鳥盤類]]などの他の恐竜のグループにも見られている。その過程においては、繊維状構造が失われ、鱗が再出現したのである<ref>{{Cite web|url=https://blog.thingswedontknow.com/2014/09/ten-things-we-dont-know-about-tyrannosaurs.html |title=Ten Things We Don't Know about Tyrannosaurs |first=Guest |last=blogger |accessdate=2021-01-05 |archivedate=2020-09-30 |archiveurl=https://web.archive.org/web/20200930075917/https://blog.thingswedontknow.com/2014/09/ten-things-we-dont-know-about-tyrannosaurs.html}}</ref>。この機序としては大型化が提唱されているが、フィル・R・ベルは羽毛のあるユウティラヌスとゴルゴサウルスおよびアルバートサウルスの体サイズが近いことを指摘した。ベルは体サイズと気候が類似することを挙げ、差異の理由が分からないとコメントした<ref>{{Cite web|url=https://www.washingtonpost.com/news/speaking-of-science/wp/2017/06/06/tyrannosaurus-rex-had-scaly-skin-and-wasnt-covered-in-feathers-a-new-study-says/ |date=2017-06-06 |archivedate=2017-06-16 |accessdate=2022-10-10 |archiveurl=https://web.archive.org/web/20170616111207/https://www.washingtonpost.com/news/speaking-of-science/wp/2017/06/06/tyrannosaurus-rex-had-scaly-skin-and-wasnt-covered-in-feathers-a-new-study-says/ |title=Tyrannosaurus rex had scaly skin and wasn’t covered in feathers, a new study says |publisher=[[ワシントン・ポスト|The Washington Post]]}}</ref>。 |
|||
=== 視界 === |
|||
[[File:Sue TRex Skull Full Frontal.JPG|thumb|upright|ティラノサウルスの[[眼窩]]は前方に向き、[[立体視]]が可能である]] |
|||
ティラノサウルスは眼窩が前側に着くことから、その眼球は前側を向き、現生の[[鷹|タカ]]よりも僅かに良い両眼視機能を持った。一般的な捕食性獣脚類は頭蓋骨の真正面に両眼視機能を持つが、ティラノサウルスは広範囲で視野が重複した。[[ジャック・ホーナー]]は、ティラノサウルス類の系統が着実に両眼視機能を向上させてきたことを指摘した。ティラノサウルス類が純粋な[[腐肉食|腐肉食動物]](スカベンジャー)であった場合、[[立体視]]をもたらす発達した奥行感覚は不要であり、この長期的な傾向が[[自然選択]]に好まれることは考えにくい<ref name="Stevens2006Binocular">{{Cite journal |last=Stevens |first=Kent A. |date=June 2006 |title=Binocular vision in theropod dinosaurs |journal=Journal of Vertebrate Paleontology |issue=2 |pages=321–330 |url=http://reocities.com/Athens/Bridge/4602/theropod_binocularvision.pdf |doi=10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 |volume=26 |s2cid=85694979 |issn=0272-4634 |url-status=dead |archiveurl=https://web.archive.org/web/20120323155306/http://reocities.com/Athens/Bridge/4602/theropod_binocularvision.pdf |archivedate=2012-03-23 }}</ref><ref name=jaffe>{{Cite journal |last=Jaffe |first=Eric |date=1 July 2006 |title=Sight for 'Saur Eyes: ''T. rex'' vision was among nature's best |journal=Science News |volume=170 |issue=1 |url=http://www.sciencenews.org/view/generic/id/7500/title/Sight_for_Saur_Eyes_%3Ci%3ET._rex%3Ci%3E_vision_was_among_natures_best |accessdate=6 October 2008 |doi=10.2307/4017288 |jstor=4017288 |pages=3–4 |archivedate=29 September 2012 |archiveurl=https://web.archive.org/web/20120929190336/http://www.sciencenews.org/view/generic/id/7500/title/Sight_for_Saur_Eyes_%3Ci%3ET._rex%3Ci%3E_vision_was_among_natures_best }}</ref>。現生の動物では、立体視は主に捕食動物に見られる。ティラノサルスと異なり、タルボサウルスは他のティラノサウルス科に典型的なより狭い頭蓋骨を持ち、眼球は主に横に面していた<ref name=savelievalifanov2005>{{Cite journal|last=Saveliev |first=Sergei V. |author2=Alifanov, Vladimir R. |year=2005 |title=A new study of the brain of the predatory dinosaur ''Tarbosaurus bataar'' (Theropoda, Tyrannosauridae) |journal=Paleontological Journal |volume=41 |issue=3 |pages=281–289 |doi=10.1134/S0031030107030070|s2cid=53529148 }}</ref>。ゴルゴサウルスの標本では、眼窩は楕円形あるいは鍵穴型というよりもむしろ円形であり、他のティラノサウルス科の属と異なる<ref name=currie2003a/>。ダスプレトサウルスでは、これは上下に高い楕円形であり、ゴルゴサウルスに見られる円形とティラノサウルスに見られる鍵穴型の中間である<ref name=holtz2004/><ref name=currie2003a/><ref name=carr1999>{{Cite journal |doi=10.1080/02724634.1999.10011161 |last=Carr |first=Thomas D. |year=1999 |title=Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria) |journal=Journal of Vertebrate Paleontology |volume=19 |issue=3 |pages=497–520 |s2cid=83744433 |url=https://zenodo.org/record/3371479 |accessdate=2019-12-02 |archivedate=2020-02-20 |archiveurl=https://web.archive.org/web/20200220183407/https://zenodo.org/record/3371479 |url-status=live }}</ref>。 |
|||
=== 顔面の感覚 === |
|||
ダスプレトサウルスと現生のワニのボーンテクスチャの比較に基づき、トーマス・D・カールらの2017年の詳細な研究でティラノサウルス科は吻部に大型で平坦な鱗を持つことが判明した<ref name="ReferenceA">{{Cite journal|last1=Carr |first1=Thomas D. |last2=Varricchio |first2=David J. |last3=Sedlmayr |first3=Jayc C. |last4=Roberts |first4=Eric M. |last5=Moore |first5=Jason R. |date=2017-03-30 |title=A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system |journal=Scientific Reports |language=en |volume=7 |doi=10.1038/srep44942 |pmid=28358353 |issn=2045-2322 |page=44942 |bibcode=2017NatSR...744942C |pmc=5372470}}</ref><ref name="Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii">{{cite journal |last1=Barker |first1=Chris |last2=Naish |first2=Darren |last3=Katsamenis |first3=Orestis |last4=Dyke |first4=Gareth |title=Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii |journal=Scientific Reports |date=June 16, 2017 |volume=7 |issue=1 |page=3749 |doi=10.1038/s41598-017-03671-3 |pmid=28623335 |pmc=5473926 |bibcode=2017NatSR...7.3749B |doi-access=free }}</ref>。鱗の中心には[[ケラチン]]質の小型のパッチが存在した。ワニにおいては、そのようなパッチは化学的刺激・熱的刺激・機械的刺激を検知可能な感覚ニューロンの束を被覆する<ref>{{Cite journal|last1=Leitch |first1=Duncan B. |last2=Catania |first2=Kenneth C. |date=2012-12-01 |title=Structure, innervation and response properties of integumentary sensory organs in crocodilians |journal=Journal of Experimental Biology |language=en |volume=215 |issue=23 |pages=4217–4230 |doi=10.1242/jeb.076836 |issn=0022-0949 |pmid=23136155 |pmc=4074209}}</ref><ref>{{Cite journal|last1=Di-Poï |first1=Nicolas |last2=Milinkovitch |first2=Michel C. |date=2013-07-02 |title=Crocodylians evolved scattered multi-sensory micro-organs |journal=EvoDevo |volume=4 |issue=1 |pages=19 |doi=10.1186/2041-9139-4-19 |pmid=23819918 |pmc=3711810 |issn=2041-9139}}</ref>。彼らは、ティラノサウルス科がおそらく顔面の鱗の下に感覚ニューロンの束を有しており、ティラノサウルス科はそれを用いて対象を特定し、[[巣]]の温度を計測し、静かに[[卵]]や幼体を持ち上げた可能性があると提唱した<ref name="ReferenceA"/>。 |
|||
=== 骨質のクレスト === |
|||
[[Image:Alioramus skull steveoc.png|thumb|明瞭な鼻骨の隆起を伴うアリオラムスの頭蓋骨]] |
|||
数多くのティラノサウルス科恐竜を含め、多数の獣脚類の頭蓋骨には骨質のクレストが見られる。モンゴルから産出した[[アリオラムス]]は、鼻骨上に卓越した5個の骨質の隆起からなる1本の列を持つ。より隆起が低いものの、同様の列は[[アパタチオサウルス]]の頭蓋骨にも存在し、ダスプレトサウルスやアルバートサウルスおよびタルボサウルスの一部の標本にも見られる<ref name=carretal2005/>。アルバートサウルスとゴルゴサウルスおよびダスプレトサウルスにおいては、涙骨上の眼の正面に卓越した角が存在する。涙骨の角はタルボサウルスとティラノサウルスには存在せず、その代わりに[[後眼窩骨]]上の眼の後方に三日月形のクレストが存在する。これらの頭部のクレストはディプレイに用いられた可能性があり、種の認識や求愛行動に用いられた可能性もある<ref name=holtz2004/>。 |
|||
=== 体温調節 === |
|||
大部分の恐竜と同様にティラノサウルスは長らく[[外温性]]と考えられていたが、1960年代後半から始まった[[恐竜ルネサンス]]の初期に[[ロバート・T・バッカー]]および[[ジョン・オストロム]]などの研究者はこの見解に対し異議を唱えた<ref name="bakker1968">{{Cite journal|last=Bakker |first=Robert T. |year=1968 |title=The superiority of dinosaurs |journal=Discovery |volume=3 |issue=2 |pages=11–12}}</ref><ref name="bakker1972">{{Cite journal|last=Bakker |first=Robert T. |year=1972 |title=Anatomical and ecological evidence of endothermy in dinosaurs |journal=Nature |volume=238 |pages=81–85 |doi=10.1038/238081a0 |bibcode=1972Natur.238...81B |issue=5359 |s2cid=4176132 }}</ref>。ティラノサウルス・レックスは[[内温性]]であるとされ、非常に活発な生態が示唆される<ref name=bakker1986>{{Cite book |last=Bakker |first=Robert T. |year=1986 |title=The Dinosaur Heresies |location=New York |publisher=Kensington Publishing |isbn=978-0-688-04287-5 |oclc=13699558 |url=https://archive.org/details/dinosaurheresies00robe }}{{Page needed|date=September 2010}}</ref>。それ以降、複数の古生物学者がティラノサウルスの体温調節能力を明らかにしようと努めてきた。若いティラノサウルス・レックスが哺乳類や鳥類と比較して高い成長率を持っていたヒストロジー的証拠は、高い[[代謝]]を有した仮説を示唆する可能性がある。哺乳類や鳥類におけるものと同様に、ティラノサウルス・レックスの成長曲線からは、本種の成長が主として未成熟個体に制限されていたことが示唆される。これは永続的に成長を遂げる他の[[脊椎動物]]の大半とは異なる<ref name="hornerpadian2004"/>。[[胴椎]]と[[脛骨]]の体温差は4 - 5℃を以下であったことが示唆される。体の中枢と末梢での温度範囲が小さいことから、古生物学者リース・バリックと地球化学者ウィリアム・シャワーズは、ティラノサウルス・レックスの体内温度は恒常的であり、外温性の爬虫類と内温性の哺乳類の中間程度の代謝を有していたとした<ref name="barrettshowers1994">{{Cite journal|first=Reese E. |last=Barrick |author2=William J. Showers |title=Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes |journal=Science |volume=265 |issue=5169 |pages=222–224 |date=July 1994 |pmid=17750663 |doi=10.1126/science.265.5169.222 |bibcode=1994Sci...265..222B |s2cid=39392327 }}</ref>。後に、数百万年過去の時代で別の大陸に生息していた[[ギガノトサウルス]]の標本においても彼らは同様の結果を発見した<ref name="barrickshowers1999">{{Cite journal |last=Barrick |first=Reese E. |author2=William J. Showers |date=October 1999 |title=Thermophysiology and biology of ''Giganotosaurus'': comparison with ''Tyrannosaurus'' |journal=Palaeontologia Electronica |volume=2 |issue=2 |url=http://palaeo-electronica.org/1999_2/gigan/issue2_99.htm |accessdate=2008-10-12 |archivedate=2011-05-17 |archiveurl=https://web.archive.org/web/20110517220816/http://palaeo-electronica.org/1999_2/gigan/issue2_99.htm }}</ref>。もしティラノサウルス・レックスに体温を恒常的に維持した証拠が無いとしても、それは外温性であったことを必ずしも意味しない。このような体温調節は現生の[[ウミガメ]]のように、大型動物ゆえの保温により説明される<ref name="paladinoetal1997">{{Cite book|editor=James O. Farlow |editor2=M. K. Brett-Surman |title=The Complete Dinosaur |publisher=Indiana University Press |location=Bloomington |year=1999 |pages=491–504 |isbn=978-0-253-21313-6 |chapter=A blueprint for giants: modeling the physiology of large dinosaurs |first1=Frank V. |last1=Paladino |first2=James R. |last2=Spotila |first3=Peter |last3=Dodson}}</ref><ref name="chinsamyhillenius2004">{{Cite book|editor=David B. Weishampel |editor2=Peter Dodson |editor3=Halszka Osmólska |title=The dinosauria |publisher=University of California Press |location=Berkeley |year=2004 |pages=643–659 |isbn=978-0-520-24209-8 |chapter=Physiology of nonavian dinosaurs |first=Anusuya |last=Chinsamy |author2=Willem J. Hillenius }}</ref><ref>{{Cite journal|last=Seymour|first=Roger S.|date=2013-07-05|title=Maximal Aerobic and Anaerobic Power Generation in Large Crocodiles versus Mammals: Implications for Dinosaur Gigantothermy|journal=PLOS ONE|volume=8|issue=7|pages=e69361|doi=10.1371/journal.pone.0069361|pmid=23861968|pmc=3702618|issn=1932-6203|bibcode=2013PLoSO...869361S|doi-access=free}}</ref>。 |
|||
== 古生態 == |
|||
===ダスプレトサウルスとゴルゴサウルスの共存 === |
|||
[[Image:Daspletosaurus FMNH.jpg|thumb|シカゴの[[フィールド自然史博物館]]にて、ダスプレトサウルスの骨格(FMNH PR308)]] |
|||
ダイナソーパーク累層において、ゴルゴサウルスはより希少なティラノサウルス亜科の種であるダスプレトサウルスと共存した。これは2属のティラノサウルス類が共存した数少ない例である。現生の捕食動物のギルドにおいて、同様の体サイズの捕食動物は競争を制限する解剖学的・行動学的・地理学的な差異により異なる[[生態的地位]](ニッチ)に分裂する。ダイナソーパーク累層のティラノサウルス科におけるニッチ分割は多くが不明である<ref name=farlowpianka2002>{{Cite journal |last=Farlow |first=James O. |author2=Pianka, Eric R. |year=2002 |title=Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs |journal=Historical Biology |volume=16 |issue=1 |pages=21–40 |doi=10.1080/0891296031000154687 |s2cid=18114585 |url=https://semanticscholar.org/paper/ccb6ab8b4949d95a9b09ef3fdbe7114b65ecb525 |accessdate=2019-12-02 |archivedate=2021-11-10 |archiveurl=https://web.archive.org/web/20211110091020/https://www.semanticscholar.org/paper/Body-Size-Overlap%2C-Habitat-Partitioning-and-Living-Farlow-Planka/ccb6ab8b4949d95a9b09ef3fdbe7114b65ecb525 }}</ref>。1970年、デイル・ラッセルは、より個体数が少なく狩りが困難である[[角竜類]]と[[曲竜類]]がダスプレトサウルスに残されていたと仮説を立てた<ref name=russell1970/>。しかし、トゥーメディスン累層から産出したダスプレトサウルスの標本 OTM 200 には、その腹部に[[消化]]されたハドロサウルス類の幼体の遺骸が保存されている<ref name=varricchio2001>{{Cite journal|last=Varricchio |first=David J. |year=2001 |title=Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts |journal=Journal of Paleontology |volume=75 |issue=2 |pages=401–406 |doi=10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2|s2cid=86113170 |issn=0022-3360}}</ref>。他のいくつかの獣脚類のグループと異なり、どちらの属も標高差による化石産出量の多寡は認められない<ref name=farlowpianka2002/>。しかし、ゴルゴサウルスはダイナソーパーク累層のような北方の層でより一般的であり、ダスプレトサウルスの種は南でより豊富である。同様のパターンは他の恐竜のグループにも見られる([[カンパニアン]]期の北アメリカ南西の[[カスモサウルス亜科]]および[[ハドロサウルス亜科]]、北アメリカ北部の[[セントロサウルス亜科]]および[[ランベオサウルス亜科]])。ホルツは、このパターンはティラノサウルス亜科・カスモサウルス亜科・ハドロサウルス亜科の間で生態的嗜好性が共有されていることを示唆しているとした。[[マーストリヒチアン]]期の末には、ティラノサウルス・レックスのようなティラノサウルス亜科、[[エドモントサウルス]]のようなハドロサウルス亜科、[[トリケラトプス]]のようなカスモサウルス亜科が北アメリカ西部に広く分布した。その一方、アルバートサウルス亜科とセントロサウルス亜科は絶滅し、ランベオサウルス亜科は衰退した<ref name=holtz2004/>。 |
|||
=== 社会的行動 === |
|||
ティラノサウルス科の間で社会的行動があった証拠も僅かに存在する。スーの標本と同じ産地では亜成体および幼体の骨格が発見・報告されており、ティラノサウルス類がある種の社会集団の中で生活していたという仮説の支持に用いられている<ref name="sue">Guinness World Records Ltd. (2003). ''2003 Guinness World Records''. pg 90.</ref>。ゴルゴサウルスには群居性の行動の証拠は存在しないが<ref name=tankecurrie1998/><ref name=currieetal2005/>、アルバートサウルスやダスプレトサウルスは群れで行動した証拠が存在する<ref>Strganac, C., Jacobs L., Polcyn M., Mateus O., Myers T., Araújo R., Fergunson K. M., Gonçalves A. O., Morais M. L., Schulp A. S., da Tavares T. S., & Salminen J. (2014). Geological Setting and Paleoecology of the Upper Cretaceous Bench 19 Marine Vertebrate Bonebed at Bentiaba, Angola. Netherlands Journal of Geosciences. 1-16.</ref>。 |
|||
ダイナソーパーク累層のダスプレトサウルスの種の若い標本 TMP 94.143.1 では、他のティラノサウルス類に起因する噛み後が顔に存在する。噛み跡は治癒しており、噛まれた後も個体が生き延びたことが示唆される。完全に成長したダイナソーパーク累層のダスプレトサウルス TMP 85.62.1 もティラノサウルス類の噛み跡を示しており、顔面への攻撃が若い個体のみに限ったものではないことが示唆される。噛み後は他の種のものである可能性もあるものの、顔面への噛み付きなど種内攻撃は捕食動物の間で非常によく見られることである。顔面への噛み付きはゴルゴサウルスやティラノサウルスといった他のティラノサウルス類だけでなく、[[シンラプトル]]や[[サウロルニトレステス]]といった他の獣脚類の属にも見られる。ダレン・ダンケとフィリップ・カリーは、噛み跡は縄張りや資源、あるいは社会集団内の支配をを巡る種内競争に起因すると仮説を立てた<ref name=tankecurrie1998/>。 |
|||
[[Image:Albertosaurus.jpg|thumb|left|アルバートサウルスのジオラマ]] |
|||
ダスプレトサウルスが社会集団として生息していた証拠は、モンタナ州のトゥーメディスン累層の[[ボーンベッド]]から得られている。ボーンベッドには3頭のダスプレトサウルスの遺骸を含んでおり、これらは大型の成体・小型の幼体・中間サイズの個体からなる。同産地には少なくとも5頭のハドロサウルス類が保存されている。地質学的証拠からは、遺骸は川の流れによって集積されたのではなく、全ての動物が同じ場所に同時に埋没したことが判明している。ハドロサウルス類の遺骸は分散しており、またティラノサウルス類の歯による多数の噛み跡が残されている。これは死亡時にダスプレトサウルスがハドロサウルス類を採餌していたことを示唆している。カリーは、確証を持つことはできないもののダスプレトサウルスが群れを形成していたことを推測した<ref name=currieetal2005/>。他の研究者は、ダスプレトサウルスや他の大型獣脚類が社会集団を形成した証拠について懐疑的である<ref name=eberthmccrea2001>{{Cite journal|last=Eberth |first=David A. |author2=McCrea, Richard T. |year=2001 |title=Were large theropods gregarious? |journal=Journal of Vertebrate Paleontology |volume=21 |issue=Supplement to Number 3 |pages=46A|doi=10.1080/02724634.2001.10010852|s2cid=220414868 }}</ref>。ブライアン・ローチとダニエル・ブリンクマンは、ダスプレトサウルスの社会相互作用は現生の[[コモドオオトカゲ]]のものに類似するものであるとした。コモドオオトカゲの群れは非協力的であり、死骸にありつき、その過程で互いに共食いすることも頻繁にある<ref name=BRDB07>{{Cite journal|last=Roach|first=Brian T.|author2=Brinkman, Daniel L. |year=2007|title=A reevaluation of cooperative pack hunting and gregariousness in ''Deinonychus antirrhopus'' and other nonavian theropod dinosaurs|journal=Bulletin of the Peabody Museum of Natural History |volume=48 |issue=1 |pages=103–138 |doi=10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 |s2cid=84175628 }}</ref>。 |
|||
バーナム・ブラウンらが発見したドライアイランドのボーンベッドには、22頭のアルバートサウルスの遺骸が保存されている。これは白亜紀の獣脚類の中で最も多くの個体が1つの場所で発見された例であり、大型獣脚類の中ではユタ州のクリーブランド=ロイド恐竜採石場の[[アロサウルス]]群集に次いで2番目に多い。この集団には非常に高齢な1頭の成体、17歳から23歳の8頭の成体、12歳から16歳で急成長段階にある7頭の亜成体、2歳から11歳の成長段階に達していない6頭の幼体が含まれる<ref name=ericksonetal2006/>。植物食性動物の遺骸がほぼ存在しないこと、およびアルバートサウルスのボーンベッドでの数多くの個体の保存状態との類似から、フィリップ・カリーはこの産地が[[カリフォルニア州]]の[[ラ・ブレア・タールピット]]のような捕食動物の罠だったのではなく、保存された全ての動物が同時に死亡したと結論した。カリーはこれを群れとしての行動の根拠とした<ref name=currie1998>{{Cite journal |last=Currie |first=Philip J. |year=1998 |title=Possible evidence of gregarious behavior in tyrannosaurids |journal=Gaia |volume=15 |pages=271–277 |url=http://www.mnhn.ul.pt/geologia/gaia/21.pdf |accessdate=2008-09-13 |archivedate=2009-03-26 |archiveurl=https://web.archive.org/web/20090326062833/http://www.mnhn.ul.pt/geologia/gaia/21.pdf }}</ref>。他の研究者は懐疑的であり、旱魃や洪水あるいは他の要因により動物たちが運搬されたと解釈した<ref name=ericksonetal2006/><ref name=eberthmccrea2001 /><ref name=roachbrinkman2007>{{Cite journal|last=Roach |first= Brian T. |author2=Brinkman, Daniel T. |year=2007 |title=A reevaluation of cooperative pack hunting and gregariousness in ''Deinonychus antirrhopus'' and other nonavian theropod dinosaurs |journal=Bulletin of the Peabody Museum of Natural History |volume=48 |issue=1 |pages=103–138 |doi=10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 |s2cid= 84175628 }}</ref>。 |
|||
一般に議論が続いているものの、少なくともいくつかのティラノサウルス科の種が社会的であったという仮説を支持する証拠は存在する。[[ブリティッシュコロンビア州]]のワピチ累層では、地元住民のアーロン・フレッドランドにより3頭のティラノサウルス科の個体に由来する足跡が発見され、リチャード・マッケラらにより記載された。足跡の調査からは、1つの足跡が作られた後に長期間放置された痕跡が認められず、3頭のティラノサウルス類の個体が集団で移動していたという仮説が支持された。さらなる調査では、これらの動物が時速6.3 - 8.4キロメートルで移動し、腰の高さは2.1 - 2.7メートルであった可能性が高いことが判明した。この層からはティラノサウルス科の異なる3属(ゴルゴサウルスとダスプレトサウルスおよびアルバートサウルス)が知られており、どの属が足痕を残したかは不明である<ref>{{Cite journal | doi=10.1371/journal.pone.0103613| pmid=25054328| pmc=4108409|title = A 'Terror of Tyrannosaurs': The First Trackways of Tyrannosaurids and Evidence of Gregariousness and Pathology in Tyrannosauridae| journal=PLOS ONE| volume=9| issue=7| pages=e103613|year = 2014|last1 = McCrea|first1 = Richard T.| last2=Buckley| first2=Lisa G.| last3=Farlow| first3=James O.| last4=Lockley| first4=Martin G.| last5=Currie| first5=Philip J.| last6=Matthews| first6=Neffra A.| last7=Pemberton| first7=S. George| bibcode=2014PLoSO...9j3613M| doi-access=free}}</ref><ref>{{cite web| url=http://www.popularmechanics.com/science/environment/tyrannosaurus-trackways-reveal-a-dinosaurs-walk-17016027 | title=Tyrannosaurus Tracks Show Dinosaur's Walk| date=2014-07-23| accessdate=2015-01-16| archivedate=2014-09-30| archiveurl=https://web.archive.org/web/20140930213639/http://www.popularmechanics.com/science/environment/tyrannosaurus-trackways-reveal-a-dinosaurs-walk-17016027 }}</ref><ref>{{cite web| url=http://phenomena.nationalgeographic.com/2014/07/23/tracks-hint-at-the-social-life-of-tyrant-dinosaurs/| title=Tracks Hint at the Social Life of Tyrant Dinosaurs| date=2014-07-23| accessdate=2015-01-16| archivedate=2015-01-08| archiveurl=https://web.archive.org/web/20150108013825/http://phenomena.nationalgeographic.com/2014/07/23/tracks-hint-at-the-social-life-of-tyrant-dinosaurs/ }}</ref>。さらなる証拠は、[[ユタ州]]南部のカイパロウィッツ累層のRainbows and Unicorns Quarryのボーンベッドにある。この化石証拠は[[テラトフォネウス]]のものとされており、2021年に記載され、他のティラノサウルス科も社会的動物であったことが示唆された。4 - 22歳に亘る異なる4 - 5頭の動物からなるこの化石からは、おそらく洪水、あるいはより低い可能性で[[藍藻]]の毒や火災や旱魃によって引き起こされた大量死事変が示唆されている。テラトフォネウスやアルバートサウルス、ティラノサウルスやダスプレトサウルスといった属のボーンベッドはティラノサウルス科に広く社会的行動が見られたことを意味しており、保存された全ての動物が短期間に死滅したという事実はティラノサウルス科の群居性行動の論拠を補強している<ref>{{Cite journal|title=Geology and taphonomy of a unique tyrannosaurid bonebed from the upper Campanian Kaiparowits Formation of southern Utah: implications for tyrannosaurid gregariousness |first1=Alan L. |last1=Titus |first2=Katja |last2=Knoll |first3=Joseph J. W. |last3=Sertich |first4=Daigo |last4=Yamamura |first5=Celina A. |last5=Suarez |first6=Ian J. |last6=Glasspool |first7=Jonathan E. |last7=Ginouves |first8=Abigail K. |last8=Lukacic |first9=Eric M. |last9=Roberts |date=April 19, 2021 |journal=PeerJ |volume=9 |pages=e11013 |doi=10.7717/peerj.11013 |pmid=33976955 |pmc=8061582 |doi-access=free}}</ref><ref>{{cite news|url=https://www.washingtonpost.com/climate-environment/2021/04/19/grand-staircase-tyrannosaur/ |title=Tyrannosaurs likely hunted in packs rather than heading out solo, scientists find |last=Eilperin |first=Juliet |date=April 19, 2021 |newspaper=The Washington Post |archiveurl=https://archive.today/20210420094932/https://www.washingtonpost.com/climate-environment/2021/04/19/grand-staircase-tyrannosaur/ |archivedate=April 20, 2021 |accessdate=September 27, 2021}}</ref><ref>{{Cite web|url=https://www.telegraph.co.uk/news/science/dinosaurs/8589113/Tyrannosaurus-Rex-hunted-in-packs.html |title=Tyrannosaurus Rex 'hunted in packs' |accessdate=2021-09-27 |archivedate=2021-09-27 |archiveurl=https://web.archive.org/web/20210927165533/https://www.telegraph.co.uk/news/science/dinosaurs/8589113/Tyrannosaurus-Rex-hunted-in-packs.html }}</ref>。 |
|||
=== 食餌 === |
|||
ティラノサウルス類の歯型は肉食恐竜の食痕として最も広く保存されており<ref name="saurornitholestes" />、角竜類やハドロサウルス類および他のティラノサウルス類から報告されている<ref name="saurornitholestes" />。既知のティラノサウルス科の骨のち約2%には歯型が保存されている<ref name="saurornitholestes">Jacobsen, A.R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.</ref>。ティラノサウルス科の歯は[[ナイフ]]のように切断する機能よりもむしろ[[肉]]を体から引き剥がす金具のように用いられた<ref name="alberto-tooth">Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: ''Mesozoic Vertebrate Life''. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.</ref>。歯の摩耗パターンからは、ティラノサウルス類の食餌には頭を振る複雑な行動が含まれていたことが示唆される<ref name="alberto-tooth"/>。 |
|||
[[File:GorgosaurusDB.jpg|thumb|left|[[パラサウロロフス]]を襲う[[テラトフォネウス]]]] |
|||
少数の研究者は、アルバートサウルスの群れでの狩猟行動について、群れの若いメンバーが獲物をより大型かつ強力であるが緩慢な成体まで誘導したと推測した<ref name=currie1998/>。幼体は成体と異なる生態を持っていた可能性があり、巨大な成体とより小型の同時代の獣脚類との間のニッチを生めていた可能性がある。同時代の小型獣脚類は、その最大の種でも成体のアルバートサウルスよりも体重が2桁小さいのである<ref name=holtz2004/>。しかし、化石記録に行動が保存されることは非常に稀であり、これらのアイディアは容易に検証が可能なものではない。フィリップ・カリーは、ダスプレトサウルスが群れを形成して狩りを行ったと思弁したが、確証を持っての主張は出来ないとした<ref name=currieetal2005/>。ゴルゴサウルスでそのような行動の証拠は発見されていない<ref name=tankecurrie1998>{{Cite journal |last=Tanke |first=Darren H. |author2=Currie, Philip J. |year=1998 |title=Head-biting behavior in theropod dinosaurs: paleopathological evidence |journal=Gaia |volume=15 |pages=167–184 |url=http://www.mnhn.ul.pt/geologia/gaia/12.pdf |accessdate=2008-09-13 |archivedate=2008-02-27 |archiveurl=https://web.archive.org/web/20080227134632/http://www.mnhn.ul.pt/geologia/gaia/12.pdf }}</ref><ref name=currieetal2005>{{Cite book|last1=Currie |first1=Philip J. |last2=Trexler |first2=David |last3=Koppelhus |first3=Eva B. |last4=Wicks |first4=Kelly |last5=Murphy |first5=Nate |year=2005 |chapter=An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA) |editor=Carpenter, Kenneth |title=The Carnivorous Dinosaurs |publisher=Indiana University Press |location=Bloomington |pages=313–324 |isbn=978-0-253-34539-4}}</ref>。 |
|||
ティラノサウルスが捕食動物であったか腐肉食動物であったかという議論は、そのロコモーションに関する議論と同程度に昔からのものである。Lambe (1917) はティラノサウルスに近縁な[[ゴルゴサウルス]]の骨格を記載し、ゴルゴサウルスの歯がほぼ摩耗していないことに基づいてゴルゴサウルスおよびティラノサウルスが純粋な腐肉食動物であったと結論した<ref name="Lambe1917Gorgosaurus">{{Cite journal|author=Lambe, L. B.|year=1917|title=The Cretaceous theropodous dinosaur ''Gorgosaurus'' |journal=Memoirs of the Geological Survey of Canada |volume=100 |pages=1–84|doi=10.4095/101672 |url=https://www.biodiversitylibrary.org/item/252624 |accessdate=2021-09-23 |archivedate=2021-09-09 |archiveurl=https://web.archive.org/web/20210909121735/https://www.biodiversitylibrary.org/item/252624 }}</ref>。獣脚類の歯は極めて急速に置換されるため、この主張は真剣には受け止められていない。ティラノサウルスの最初の発見以降大半の研究者はティラノサウルスが捕食動物であったことに同意しているが、現生の捕食動物のように機会があれば喜んで他の捕食者の獲物を盗んだり死肉を漁ったりしたと思われる<ref name="FarlowHoltz2002FossilRecordPredation">{{Cite web|editor=Kowalewski, M.|editor2=Kelley, P.H|title=The Fossil Record of Predation |year=2002 |series=The Paleontological Society Papers |volume=8 |pages=251–266 |url=http://www.yale.edu/ypmip/predation/Chapter_09.pdf |author1=Farlow, J. O. |author2=Holtz, T. R. |archiveurl=https://web.archive.org/web/20081031093048/http://www.yale.edu/ypmip/predation/Chapter_09.pdf |archivedate=2008-10-31 |accessdate=2022-10-11}}</ref><ref name="dinocards2">Dorey, M. (1997). Tyrannosaurus. ''Dinosaur Cards''. Orbis Publishing Ltd. D36045907.</ref>。 |
|||
[[ジャック・ホーナー]]は、ティラノサウルスが専ら腐肉食動物であり、一切の狩りを行わなかったと考えた<ref name="hornerlessem1993" /><ref name="Horner1994SteakKnives">{{Cite journal|author=Horner, J.R.|year=1994|title=Steak knives, beady eyes, and tiny little arms (a portrait of ''Tyrannosaurus'' as a scavenger) |journal=The Paleontological Society Special Publication |volume=7 |pages=157–164 |doi=10.1017/S2475262200009497}}</ref><ref name="BBC2003TrexOnTrial">{{Cite news|url=http://news.bbc.co.uk/1/hi/sci/tech/3112527.stm |title=T. rex goes on trial |author=Amos, J. |publisher=BBC |date=31 July 2003 |accessdate=12 October 2008 |archivedate=13 March 2012 |archiveurl=https://web.archive.org/web/20120313130020/http://news.bbc.co.uk/1/hi/sci/tech/3112527.stm }}</ref>。ホーナーは純粋腐肉食動物仮説を支持する複数の主張を紹介している。大型の[[嗅球]]と嗅神経の存在からは、遠距離の死骸を嗅ぎ付けるための高度に発達した嗅覚が示唆される。歯は骨を砕くことができ、それゆえ通常は最も栄養分の少ない部位である死体の残りから可能な限りの食料([[骨髄]])を抜き取ることが可能だった。また、ティラノサウルスは走るのではなく歩いていたことが証拠から示唆されているが、少なくともその獲物となりうる動物は俊敏に動くことが出来た<ref name="Horner1994SteakKnives" /><ref name="dinodictionary"/>。 |
|||
他の証拠からは、ティラノサウルスの狩猟行動が示唆されている。ティラノサウルス類の眼窩は目が前を向くように位置しており、良好な[[立体視]]が可能であった。ティラノサウルス類が負わせた負傷が[[トリケラトプス]]やハドロサウルス類の骨格に認められており、それらは最初の攻撃を生き延びたように見える<ref name="erickson1996">{{cite journal | author1 = Erickson G. M. | author2 = Olson K. H. | year = 1996 | title = Bite marks attributable to ''Tyrannosaurus rex'': preliminary description and implications | url = https://zenodo.org/record/3732430 | journal = Journal of Vertebrate Paleontology | volume = 16 | issue = 1 | pages = 175–178 | doi = 10.1080/02724634.1996.10011297 | accessdate = 2020-09-12 | archivedate = 2021-10-19 | archiveurl = https://web.archive.org/web/20211019153802/https://zenodo.org/record/3732430 | url-status = live }}</ref><ref name=carpenter1998>{{Cite journal|last=Carpenter |first=K. |year=1998 |title=Evidence of predatory behavior by theropod dinosaurs |journal=Gaia |volume=15 |pages=135–144 |url=http://vertpaleo.org/publications/jvp/15-576-591.cfm|accessdate=5 December 2007 |archiveurl=https://web.archive.org/web/20071117132451/http://vertpaleo.org/publications/jvp/15-576-591.cfm <!--Added by H3llBot--> |archivedate=17 November 2007}}</ref><ref name="fowler2006">{{Cite journal|first=Denver W. |last= Fowler |author2=Robert M. Sullivan |year=2006 |title=A ceratopsid pelvis with toothmarks from the Upper Cretaceous Kirtland Formation, New Mexico: evidence of late Campanian tyrannosaurid feeding behavior |journal=New Mexico Museum of Natural History and Science Bulletin |volume=35 |pages=127–130}}</ref>。ティラノサウルスが腐肉食動物ならば、北アメリカからユーラシアにかけての上部白亜系に別の恐竜が頂点捕食者の地位に立っていなければならないと主張する研究者もいる。頂点捕食者の獲物は大型の[[周飾頭亜目|周飾頭類]]と[[鳥脚類]]であった。他のティラノサウルス科はティラノサウルスと数多くの特徴を共有するため、ティラノサウルスを除外した場合、頂点捕食者の候補として残るのは小型である[[ドロマエオサウルス類]]のみである。このことから、腐肉食動物仮説を信じる者は、ティラノサウルス類がその巨体と力で以て小型動物から獲物を盗むことが可能だったと提唱した<ref name="dinodictionary">Walters, M., Paker, J. (1995). Dictionary of Prehistoric Life. Claremont Books. {{ISBN2|1-85471-648-4}}.</ref>。 |
|||
==== 共食い ==== |
|||
ティラノサウルス科が少なくとも時折共食いしていたことを強く示唆する証拠もある。ティラノサウルス自体には1つの標本において足・[[上腕骨]]・[[中足骨]]に歯型が見られるものがあり、これに基づいて少なくとも死骸を漁るために共食いが行われていたことが示唆される<ref name="HornerCurrie_et.al._2010_Trex_cannibalism">{{Cite journal |last1=Longrich |first1=N. R. |last2=Horner |first2=J. R. |last3=Erickson |first3=G. M. |last4=Currie |first4=P. J. |year=2010 |title=Cannibalism in ''Tyrannosaurus rex'' |journal=PLOS ONE |volume=5 |issue=10 |page=e13419 |doi=10.1371/journal.pone.0013419 |pmc=2955550 |pmid=20976177|bibcode=2010PLoSO...513419L |doi-access=free }}</ref>。フルートランド層およびキルトランド層(共にカンパニアン期)産の化石、およびオジョ・アラモ層(マーストリヒチアン期)産の化石からは、San Juan 盆地の様々なティラノサウルス科で共食いが起きていたことが示唆される。標本から集められた証拠に基づくと、ティラノサウルス科が同種の動物の共食いを伴う日和見的な摂食行動をとっていた<ref>{{Cite web|url=https://www.researchgate.net/publication/348002335 |title=NEW EVIDENCE FOR CANNIBALISM IN TYRANNOSAURID DINOSAURS FROM THE UPPER CRETACEOUS (CAMPANIAN/MAASTRICHTIAN) SAN JUAN BASIN OF NEW MEXICO |website=ResearchGate |accessdate=2021-01-06 |archivedate=2021-11-10 |archiveurl=https://web.archive.org/web/20211110091047/https://www.researchgate.net/publication/348002335_NEW_EVIDENCE_FOR_CANNIBALISM_IN_TYRANNOSAURID_DINOSAURS_FROM_THE_UPPER_CRETACEOUS_CAMPANIANMAASTRICHTIAN_SAN_JUAN_BASIN_OF_NEW_MEXICO }}</ref>。 |
|||
===分布 === |
|||
[[Image:Tyrannosauroid fossil localities map.png|thumb|right|450px|ティラノサウルス上科の化石の分布。ティラノサウルス科は青色で示されている。]] |
|||
初期のティラノサウルス上科が[[北半球]]の全ての大陸で発見されている一方、ティラノサウルス科の化石は北アメリカとアジアでしか発見されていない。南半球で産出した断片的な化石がティラノサウルス科のものとして報告されることは複数回あったが、これらは[[アベリサウルス科]]の化石の誤同定である<ref name="ageofdinosaursabelisaurus">{{Cite book|chapter=Abelisaurus|author1=Dodson, Peter |author2=Britt, Brooks |author3=Carpenter, Kenneth |author4=Forster, Catherine A. |author5=Gillette, David D. |author6=Norell, Mark A. |author7=Olshevsky, George |author8=Parrish, J. Michael |author9= Weishampel, David B. |name-list-style=amp |title=Age of Dinosaurs|publisher=Publications International, LTD|page=105|isbn=978-0-7853-0443-2|year=1993}}</ref>。白亜紀の中頃の化石記録がアジア・北アメリカの両地域で乏しいためティラノサウルス科の起源となった時代と場所は不明なままであるが、ティラノサウルス科のものとして確認された最初期のものは[[カンパニアン]]期の北アメリカ西部に生息していた<ref name=holtz2004/>。 |
|||
北アメリカ東部でティラノサウルス科の化石は発見されていないが、[[ドリプトサウルス]]や[[アパラチオサウルス]]といったより基盤的なティラノサウルス上科の化石は回収されている。これらは白亜紀の終わりまで持続しており、このことから白亜紀の中頃に[[西部内陸海路]]によって北アメリカが分裂した後、西部でティラノサウルス科が進化したことが示唆される<ref name=carretal2005/>。ティラノサウルス科の化石は[[アラスカ州]]からも発見されており、これが北アメリカとアジアに分散したルートであった可能性がある<ref name=fiorillogangloff2000>{{Cite journal|last=Fiorillo |first=Anthony R. |author2=Gangloff, Roland A. |year=2000 |title=Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on arctic dinosaur paleoecology |journal=Journal of Vertebrate Paleontology |volume=20 |issue=4 |pages=675–682 |doi=10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2|s2cid=130766946 |issn=0272-4634}}</ref>。ある系統解析ではアリオラムスとタルボサウルスは近縁な位置に置かれており、本科において固有のアジアの系統を形成した<ref name=currieetal2003/>。後に[[キアンゾウサウルス]]の発見およびアリオラムス族の記載によりこれは反証された<ref name="Junchang2014"/>。2015年、[[日本]]の[[福井県立恐竜博物館]]は[[長崎県]][[長崎市]]の長崎半島から産出したティラノサウルス科の歯化石を報告した。これは8100万年前([[カンパニアン]]期)のもので、日本国内初の大型ティラノサウルス類の歯は化石として発表された<ref>{{Cite web|url=https://www.dinosaur.pref.fukui.jp/research/201507Nagasaki/ |date=2015-07-14 |accessdate=2022-10-06 |publisher=[[福井県立恐竜博物館]] |title=国内初のティラノサウルス科大型種の歯の化石について}}</ref>。 |
|||
[[Image:TarbosaurusP1050352.jpg|thumb|left|タルボサウルスの骨格キャスト]] |
|||
ティラノサウルス亜科はより広範囲に分布した。アルバートサウルス亜科はアジアで発見されておらず、アジアにはタルボサウルスやズケンティラヌスおよびアリオラムス族(アリオラムスやキアンゾウサウルス)といったティラノサウルス亜科が生息していた。ティラノサウルス亜科とアルバートサウルス亜科は共にカンパニアン期 - [[マーストリヒチアン]]期に北アメリカに生息しており、ダスプレトサウルスのようなティラノサウルス亜科は西部内陸中に分布した一方、アルバートサウルス亜科のアルバートサウルスやゴルゴサウルスは大陸の北西部のみでしか知られていない<ref>{{Cite journal|title=The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics|author=Thomas R. Holtz Jr.|journal=Journal of Paleontology|volume=68|issue=5|year=1994|pages=1100–1117|jstor=1306180|doi=10.1017/S0022336000026706|s2cid=129684676 }}</ref>。 |
|||
後期マーストリヒチアン期までにアルバートサウルス亜科は絶滅し、ティラノサウルス亜科が[[サスカチュワン州]]から[[テキサス州]]まで分布した。このパターンは他の北アメリカの恐竜にも当てはまっている。カンパニアン期から前期マーストリヒチアン期にかけて、北西部では[[ランベオサウルス亜科]]の[[鳥脚類]]と[[セントロサウルス亜科]]の[[角竜]]が繁栄し、南部では[[ハドロサウルス亜科]]と[[カスモサウルス亜科]]が繁栄した。白亜紀末ではセントロサウルス亜科は発見されておらず、ランベオサウルス亜科も数を減らしていた一方、ハドロサウルス亜科とカスモサウルス亜科は西部内陸中で繁栄した<ref name=holtz2004/>。2016年にスティーヴ・ブルサッテらは、北アメリカ西部の大部分の他のティラノサウルス科の絶滅にティラノサウルス自体が部分的に関与していたことを示唆した。この研究では、ティラノサウルスが北アメリカで進化したのではなく(おそらくタルボサウルスの子孫として)アジアから渡り、他のティラノサウルス科と競合して生存競争に勝利したことが示唆されている。この仮説は、ティラノサウルスが知られている範囲で他種のティラノサウルス科がほとんど発見されていない事実からも支持されている<ref>{{Cite web|url=https://www.livescience.com/53877-t-rex-was-invasive-species.html|title=T. Rex Was Likely an Invasive Species |first=Laura Geggel 29 |last=February 2016 |website=livescience.com |date=29 February 2016 |accessdate=5 January 2021 |archivedate=14 July 2019 |archiveurl=https://web.archive.org/web/20190714170020/https://www.livescience.com/53877-t-rex-was-invasive-species.html |url-status=live}}</ref>。 |
|||
== 属の時系列 == |
|||
<timeline> |
|||
ImageSize = width:800px height:auto barincrement:15px |
|||
PlotArea = left:10px bottom:50px top:10px right:10px |
|||
Period = from:-100.5 till:-60 |
|||
TimeAxis = orientation:horizontal |
|||
ScaleMajor = unit:year increment:10 start:-100 |
|||
ScaleMinor = unit:year increment:1 start:-100 |
|||
TimeAxis = orientation:hor |
|||
AlignBars = justify |
|||
Colors = |
|||
#legends |
|||
id:CAR value:claret |
|||
id:ANK value:rgb(0.4,0.3,0.196) |
|||
id:HER value:teal |
|||
id:HAD value:green |
|||
id:OMN value:blue |
|||
id:black value:black |
|||
id:white value:white |
|||
id:latecretaceous value:rgb(0.74,0.82,0.37) |
|||
BarData= |
|||
bar:eratop |
|||
bar:space |
|||
bar:periodtop |
|||
bar:space |
|||
bar:NAM1 |
|||
bar:NAM2 |
|||
bar:NAM3 |
|||
bar:NAM4 |
|||
bar:NAM5 |
|||
bar:NAM6 |
|||
bar:NAM7 |
|||
bar:NAM8 |
|||
bar:NAM9 |
|||
bar:NAM10 |
|||
bar:NAM11 |
|||
bar:NAM12 |
|||
bar:NAM13 |
|||
bar:space |
|||
bar:period |
|||
bar:space |
|||
bar:era |
|||
PlotData= |
|||
align:center textcolor:black fontsize:M mark:(line,black) width:25 |
|||
shift:(7,-4) |
|||
bar:periodtop |
|||
from: -100.5 till: -93.9 color:latecretaceous text:[[セノマニアン|Cenomanian]] |
|||
from: -93.9 till: -89.8 color:latecretaceous text:[[チューロニアン|Turonian]] |
|||
from: -89.8 till: -86.3 color:latecretaceous text:[[コニアシアン|Coniac.]] |
|||
from: -86.3 till: -83.6 color:latecretaceous text:[[サントニアン|Sant.]] |
|||
from: -83.6 till: -72.1 color:latecretaceous text:[[カンパニアン|Campanian]] |
|||
from: -72.1 till: -66 color:latecretaceous text:[[マーストリヒチアン|Maastrichtian]] |
|||
bar:eratop |
|||
from: -100.5 till: -66 color:latecretaceous text:[[後期白亜紀|Late Cretaceous]] |
|||
PlotData= |
|||
align:left fontsize:M mark:(line,white) width:5 anchor:till align:left |
|||
color:ANK bar:NAM1 from:-80.6 till:-79.9 text:[[リトロナクス|Lythronax]] |
|||
color:ANK bar:NAM2 from:-78.5 till:-78.0 text:[[ディナモテラー|Dynamoterror]] |
|||
color:ANK bar:NAM3 from:-77 till:-76 text:[[テラトフォネウス|Teratophoneus]] |
|||
color:ANK bar:NAM4 from:-77 till:-74 text:[[ダスプレトサウルス|Daspletosaurus]] |
|||
color:OMN bar:NAM5 from:-76.6 till:-75.1 text:[[ゴルゴサウルス|Gorgosaurus]] |
|||
color:ANK bar:NAM6 from:-74.84 till:-74.26 text:[[ビスタヒエヴェルソル|Bistahieversor]] |
|||
color:ANK bar:NAM7 from:-73.6 till:-73.4 text:[[ズケンティラヌス|Zhuchengtyrannus]] |
|||
color:OMN bar:NAM8 from:-71 till:-68 text:[[アルバートサウルス|Albertosaurus]] |
|||
color:ANK bar:NAM9 from:-70.1 till:-69.9 text:[[アリオラムス|Alioramus]] |
|||
color:ANK bar:NAM10 from:-70.1 till:-69.9 text:[[タルボサウルス|Tarbosaurus]] |
|||
color:ANK bar:NAM11 from:-69.1 till:-69.2 text:[[ナヌークサウルス|Nanuqsaurus]] |
|||
color:ANK bar:NAM13 from:-68 till:-66 text:[[ティラノサウルス|Tyrannosaurus]] |
|||
PlotData= |
|||
align:center textcolor:black fontsize:M mark:(line,black) width:25 |
|||
bar:period |
|||
from: -100.5 till: -93.9 color:latecretaceous text:[[セノマニアン|Cenomanian]] |
|||
from: -93.9 till: -89.8 color:latecretaceous text:[[チューロニアン|Turonian]] |
|||
from: -89.8 till: -86.3 color:latecretaceous text:[[コニアシアン|Coniac.]] |
|||
from: -86.3 till: -83.6 color:latecretaceous text:[[サントニアン|Sant.]] |
|||
from: -83.6 till: -72.1 color:latecretaceous text:[[カンパニアン|Campanian]] |
|||
from: -72.1 till: -66 color:latecretaceous text:[[マーストリヒチアン|Maastrichtian]] |
|||
bar:era |
|||
from: -100.5 till: -66 color:latecretaceous text:[[後期白亜紀|Late Cretaceous]] |
|||
</timeline> |
|||
{{Clear}} |
|||
== |
== 出典 == |
||
{{Reflist|2}} |
|||
*[[ティラノサウルス]] |
|||
*[[タルボサウルス]] |
|||
*[[アルバートサウルス]] |
|||
*[[ダスプレトサウルス]] |
|||
*[[シャモティラヌス]] |
|||
*[[ナノティラヌス]] |
|||
*[[ラプトレックス]] |
|||
*[[ディロング]] |
|||
*[[恐竜]] |
|||
{{Normdaten}} |
|||
{{DEFAULTSORT:ていらのさうるすか}} |
{{DEFAULTSORT:ていらのさうるすか}} |
||
[[Category:ティラノサウルス科|*]] |
[[Category:ティラノサウルス科|*]] |
||
2022年10月31日 (月) 09:09時点における版
| ティラノサウルス科 | ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| ||||||||||||||||||||||||||||||||||||||||||
| 地質時代 | ||||||||||||||||||||||||||||||||||||||||||
| 後期白亜紀 | ||||||||||||||||||||||||||||||||||||||||||
| 分類 | ||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||
| 学名 | ||||||||||||||||||||||||||||||||||||||||||
| Tyrannosauridae Osborn, 1905 | ||||||||||||||||||||||||||||||||||||||||||
| 亜科、属 | ||||||||||||||||||||||||||||||||||||||||||
ティラノサウルス科 (Tyrannosauridae) は、ティラノサウルス上科に属する獣脚類の恐竜の科。同上科の恐竜の中では大型から超大型であり、後期白亜紀の北アメリカ大陸とアジアに生息した。解剖学的特徴は共通性が高いが、幼体やより小型の属種の体は華奢で、大型の属種の成体は頑強な体格を有する[1]。
より小型の祖先の子孫であるにも拘わらず、ティラノサウルス科の恐竜はほぼ常時各々の生態系において最大の捕食動物であり、食物連鎖の頂点に位置していた。最大の種 Tyrannosaurus rex で、これは既知の陸上捕食動物では最大かつ最も頑強であり、全長は12.3メートルを超え[2]、体重は8.4 - 14トンと推定されている[2][3][4]。二足歩行性であり、顎にはD字型の断面を持つ歯がU字型に配列し、吻部の先端は丸みを帯びる。長い後肢からは他の大型獣脚類と比較して移動速度の向上が示唆されるが、その一方で前肢は手としての機能を保持しつつも退化しており、指も退縮して2本になっている[1]。
発見の歴史

ティラノサウルス科の最初の化石が発見されたのはカナダ地質調査所による遠征の際であり、その際には無数の離散した歯が発見された。これらの明らかな恐竜の歯は1856年にジョセフ・ライディが Deinodon(「怖ろしい歯」)として命名した。ティラノサウルス科の最初の良好な標本はアルバータ州のホースシューキャニオン累層で発見されたものであり、部分的な骨格を伴うほぼ完全な頭蓋骨から構成されていた。これらの化石は1876年にエドワード・ドリンカー・コープが最初に研究し、東方のティラノサウルス上科の属ドリプトサウルスの種のものであると考えた。1905年、ヘンリー・フェアフィールド・オズボーンはアルバータ州の化石をドリプトサウルスと顕著に異なるものであると認識し、Albertosaurus sarcophagus(「肉を食べるアルバータのトカゲ」)と命名した[5]。1892年にコープは他のティラノサウルス類の骨を記載しており、記載された単離した椎骨はManospondylus gigas と命名された。この発見は100年以上に亘って見落とされており、やがて本標本が実際にはTyrannosaurus rex のものであり、さらに当該の学名に対して先取権を持つことが判明すると、2000年代初頭に議論を呼んだ[6]。

1905年にアルバートサウルスを命名する論文を発表したオズボーンは、同年にさらなるティラノサウルス類の2標本 を記載した。これらの標本はアメリカ自然史博物館が1902年に行ったバーナム・ブラウン率いる遠征においてモンタナ州とワイオミング州で発見されたものであり、オズボーンは当初これらの標本を別種のものであると考えた。彼は1つの標本に Dynamosaurus imperiosus(「力のトカゲの皇帝」)、もう1つの標本に Tyrannosaurus rex(「暴君トカゲの王」)と命名した。1年後、オズボーンはこれらの2標本が同じ種に由来するものであると認識した。最初に発見されたのはディナモサウルスの方であったが、ティラノサウルスという名前が原記載において1ページ先に記載されていたため、ティラノサウルスの名前が用いられるようになった[7]。
バーナム・ブラウンはアルバータ州からさらなるティラノサウルス科標本を蒐集しており、当該グループに特徴的な2本指で短い前肢を保存したもの(1914年にローレンス・ラムが Gorgosaurus libratus 「均衡のとれた熾烈なトカゲ」として命名)もその中には含まれていた。ゴルゴサウルスに関わる2つ目の重大な発見は1942年のもので、保存状態が良好であるが異様に小型の完全な頭蓋骨が発見された。当該標本は第二次世界大戦後にチャールズ・W・ギルモアが研究し、Gorgosaurus lancesnis と命名した[5]。1988年にはロバート・T・バッカーとフィリップ・J・カリーおよびマイケル・ウィリアムズがこの頭蓋骨を再研究し、新属 Nanotyrannus に分類した[8]。また、1946年にソビエト連邦の古生物学者たちはモンゴル国への遠征を開始し、アジア初のティラノサウルス類の化石を発見した。1955年に Evgeny Maleev はモンゴルの化石をティラノサウルスおよびゴルゴサウルス、そして1つの新属 Tarbosaurus(「怖ろしいトカゲ」)と命名した。後の研究で、Maleev のティラノサウルス類の種は実際には成長段階の異なるタルボサウルスの種であることが判明した。後にモンゴルで発見された第二の種は1976年にセルゲイ・クルザノフが記載・命名した Alioramus remotus(「隔たった異なる枝」)であった。ただし、アリオラムスが真のティラノサウルス科であるか、あるいはより基盤的なティラノサウルス類であるかについては、いまだ議論がある[9][5]。
説明

ティラノサウルス科はいずれも大型の動物であり、全ての種が体重1トンを超える[10]。アリオラムスの1個の標本は全長5 - 6メートルの個体と推定されるものも発見されているが[9]、これを未成熟個体と考える専門家もいる[10][11]。アルバートサウルス、ゴルゴサウルス、ダスプレトサウルスはいずれも全長8 - 10メートルであり[12]、タルボサウルスは12メートルに達する[13]。ティラノサウルスの最大級の標本の一つであるスーは全長12.3メートルに達する[2]。
頭蓋骨と歯列


_of_Daspletosaurus_horneri.jpg)

部分的な頭蓋骨だけが発見されているアリオラムスを除いて全ての属で完全な頭蓋骨が得られているため、ティラノサウルス科の頭蓋骨の解剖学はよく理解されている[14]。ティラノサウルス、タルボサウルス、ダスプレトサウルスの頭蓋骨は長さ1メートルを超える[12]。成体のティラノサウルス科は高く頑強な頭蓋骨を持ち、数多くの骨が癒合して強度を増している。同時に、数多くの頭蓋骨の間に存在する空洞のチャンバーや、それらの骨の間に位置する大型の開口部(窓)は、頭蓋骨の軽量化に寄与した。高い前上顎骨や癒合した鼻骨など、ティラノサウルス科の頭蓋骨に見られる数多くの特徴は直近の祖先にも見ることが出来る[10]。
ティラノサウルス科の頭蓋骨には数多くの固有の形質がある。具体的には、卓越した矢状稜を伴う癒合した頭頂骨が挙げられる。この稜は矢状縫合に沿って縦方向に走り、頭蓋天井の2つの上側頭窓を隔てる。これらの窓の後側では、ティラノサウルス科は特徴的に高い項部のクレストを持ち、このクレストは縦方向よりもむしろ横断面に沿って頭頂骨上を走る。このクレストはティラノサウルス、タルボサウルス、アリオラムスにおいて発達する。アルバートサウルス、ダスプレトサウルス、ゴルゴサウルスは涙骨上に目の前に高いクレストを持つが、タルボサウルスとティラノサウルスは極端に厚みを帯びた後眼窩骨が三日月形のクレストを目の後ろで形成する。アリオラムスはその吻部の最上部に1列に配列した6個の骨質のクレストを持つ。ダスプレトサウルスとタルボサウルスではより低いクレストが報告された標本があり、さらに基盤的なティラノサウルス上科であるアパラチオサウルスとも共通する[11][15]。吻部と頭蓋骨の他の部位にも無数の孔が存在する。D. horneri を記載した2017年の研究によると、ワニやティラノサウルス科に見られる多列の神経血管孔は触覚や鱗状の外皮にも相関がある[16]。


ティラノサウルス科はティラノサウルス上科の祖先と同様に異歯性を持ち、前上顎骨歯は断面がD字型で、他の歯よりも小型である。最初期のティラノサウルス上科や他の獣脚類と異なり、成熟したティラノサウルス科の上顎骨歯と歯骨歯はブレード状ではなく極端に厚く、また断面は丸みを帯びており、いくつかの種では鋸歯が減少している[10]。歯の本数は種のうちで一定しており、大型の種では小型の種よりも少ない傾向がある。例えば、アリオラムスは76 - 78本、ティラノサウルスは54 - 60本の歯を持つ[17]。
ウィリアム・エイブラーは2001年に、アルバートサウルスの歯の鋸歯が歯の割れ目に類似し、膨大部と呼称される丸い空洞に帰着することを観察した[18]。ティラノサウルス科の歯は体から肉を引っ張るための留め具として用いられ、そのためティラノサウルス類が肉片を引き剥がす際の張力によって割れ目のような鋸歯が歯の一面に広がった可能性がある[18]。しかし、膨大部があれば、これらの力をより大きな表面積に分散させ、負担のかかった歯が損傷するリスクを軽減できた[18]。空洞に帰着する切れ込みの存在は、人間の工学とも相等しい。エイブラーの説明によると、ギターメーカーは空洞に帰着する切れ込みを入れることで柔軟性と剛性を交互に木材に与えるとのことである[18]。また航空機の表面の保護においては、ドリルで膨大部様構造を作り、素材の亀裂の伝播を防ぐ方法が用いられている[18]。エイブラーは切れ込みとドリルによる穿孔を伴うアクリル樹脂のバーが規則的に切り込みを入れただけのものに比べて25%以上強い強度を持つことを実証した[18]。ティラノサウルス類や他の獣脚類と異なり、植竜類やディメトロドンなどの太古の捕食動物は摂食の力を受けても歯の切れ目状の鋸歯が広がらないようにする適応を遂げなかった[18]。
体骨格

頭蓋骨は厚いS字型の頸部の先端に位置しており、長く重厚な尾は頭部と胴部に対するカウンターウエイトとして機能し、重心は臀部に位置する。ティラノサウルス科はプロポーション的に非常に小型の2本指の前肢で知られているが、痕跡器官となった第III指が残る場合もある[10][19]。

ティラノサウルス科は後肢で歩行するため、その骨は頑強である。前肢とは対照的に、大半の他のどの獣脚類と比較しても後肢は体サイズに対して長い。幼体やいくつかの小型の成体では、基盤的ティラノサウルス上科と同様に、脛骨が大腿骨よりも長く、オルニトミモサウルス類と同様に脚の速い恐竜の形質を示す。より大型の成体は動作の緩慢な動物の後肢のプロポーション形質を有しているが、アベリサウルス科やカルノサウルス類のような他の大型獣脚類に見られるほどではない。ティラノサウルス科の第III中足骨は第IIおよび第IV中足骨に挟まれて細くなっており、アークトメタターサルとして知られる構造をなす[10]。
アークトメタターサルが最初に進化した時代は不明である。ディロングのような最初期のティラノサウルス上科にアークトメタターサルは存在しないが[20]、より新しい時代のアパラチオサウルスには見られる[15]。この構造はトロオドン科やオルニトミモサウルス類およびカエグナトゥス科を特徴づけるが[21]、最初期のティラノサウルス上科に存在しないことから当該の形質が収斂進化によるものであることが示唆される[20]。
分類
Deinodontidae という名前は1866年にエドワード・ドリンカー・コープがこの科について作り[22]、1960年代まで新たな名前 Tyrannosauridae の立ち位置で用いられた[23]。Deinodontidae のタイプ属は Deinodon で、モンタナ州産の単離した歯から命名された[24]。しかし、1970年の北アメリカのティラノサウルス類のレビューにおいて、デイル・ラッセルは Deinodon が有効な分類群でないと結論し、Deinodontidae の代わりに Tyrannosauridae の名前を用い、国際動物命名規約と一致すると主張した[12]。このため、ティラノサウルス科という名前が現在の専門家から好まれている[5]。
Tyrannosaurus は1905年にヘンリー・フェアフィールド・オズボーンが命名し、これに伴ってティラノサウルス科も命名された[25]。この名前は古代ギリシア語の τυραννος (tyrannos)(「暴君」)および σαυρος (sauros)(「トカゲ」)に由来する。接尾詞 -idae は動物の科の名前に一般に付されるものであり、複数名詞を示すギリシア語の接尾詞 -ιδαι -idai に由来する[26]。
分類法

ティラノサウルス科はリンネ式階層分類体系における科であり、ティラノサウルス上科と獣脚亜目の内部に置かれる。
ティラノサウルス科は2つの亜科に細分される。アルバートサウルス亜科は北アメリカの属であるアルバートサウルスとゴルゴサウルスを含み、ティラノサウルス亜科はダスプレトサウルス、テラトフォネウス、ビスタヒエヴェルソル、タルボサウルス、ナヌークサウルス、ズケンティラヌス、ティラノサウルスを含む[27]。Gorgosaurus libratus をアルバートサウルス属に、Tarbosaurus bataar をティラノサウルス属に含む研究者もいるが[15][5][28]、分かれた属としてゴルゴサウルスとタルボサウルスを維持することを好む研究者もいる[10][11]。アルバートサウルス亜科はティラノサウルス亜科と比較してより細身の体格、低い頭蓋骨、プロポーション的に長い脛骨を特徴に持つ[10]。ティラノサウルス亜科では、頭頂骨の矢状クレストが前頭骨上に続く[11]。2014年には、Lü Junchang et al. がティラノサウルス科内の族としてアリオラムスとキアンゾウサウルスを含むアリオラムス族を記載した。彼らの系統解析では本族はティラノサウルス亜科の基底に置かれることが示唆された[29][30]。他にも George Olshevsky や Tracy Ford といった研究者が様々なティラノサウルス科恐竜の組み合わせでティラノサウルス亜科の他の下位分類や族を作成したが[31][32]、これらは系統学的に定義されておらず、またこれらを構成する属は他の属や種のシノニムと考えられている[17]。
さらなる亜科はより断片的な属にちなんで命名されており、アウブリソドン亜科とデイノドン亜科が挙げられる。しかし、アウブリソドンとデイノドンは通例疑問名と考えられており、そのため彼らや彼らのエポニムの亜科はティラノサウルス科の分類から通常除外される。さらなるティラノサウルス科として、当初はより基盤的なティラノサウルス上科と考えられていたが、タルボサウルスに類似するティラノサウルス亜科の幼体を代表する可能性が高いラプトレックスがいる。しかし、本属は幼体の標本しか知られていないため、これも疑問名と考えられている[33]。
系統
古脊椎動物学において系統分類学が出現し、ティラノサウルス科は複数の明白な定義を与えられている。最初の定義はポール・セレノによる1998年のもので、アレクトロサウルスやアウブリソドンあるいはナノティラヌスよりもティラノサウルスに近縁な全てのティラノサウルス上科が本科に含まれた[34]。しかし、ナノティラヌスはよくティラノサウルスの幼体と考えられており、アウブリソドンも疑問名であるため、系統群の定義には不適である[10]。
2001年、トーマス・R・ホルツ・ジュニアはティラノサウルス科の分岐分析を発表した[35]。彼は、鋸歯の無い前上顎骨歯で特徴づけられる原始的なアウブリソドン亜科と、ティラノサウルス亜科の2亜科が存在すると結論した[35]。アウブリソドン亜科にはアウブリソドンの他に "Kirtland Aublysodon" とアレクトロサウルスが含まれる[35]。また、ホルツはシャモティラヌスがティラノサウルス科の共有派生形質を複数示すことを発見したが、ティラノサウルス科には置かなかった[35]。
同誌で後述された記述にて、彼はティラノサウルス科を「ティラノサウルスとアウブリソドンの最も近い共通祖先の全ての子孫」として定義することを提唱した[35]。また、彼はポール・セレノのものをはじめ他の研究者が過去に提唱した定義を批判した[35]。ホルツはナノティラヌスがティラノサウルス・レックスの幼体の誤同定であると考えていたため、セレノが提唱した定義ではティラノサウルス科はティラノサウルス属の下位分類になるとしたのである[35]。さらに、セレノの提案した定義ではティラノサウルス亜科もティラノサウルスのみに限定されるとした[35]。
2003年にクリストファー・ブロチュはアルバートサウルス、アレクトロサウルス、アリオラムス、ダスプレトサウルス、ゴルゴサウルス、タルボサウルスおよびティラノサウルスを定義に加えた[36]。ホルツは2004年に系統群を再定義し、彼の分析で確固たる位置に置かれなかったアリオラムスとアレクトロサウルスを除いて上記の全てを指示子として用いた。しかし、彼は同じ論文において、エオティラヌスよりもティラノサウルスに近縁な全ての獣脚類を含むものとして別の定義も提供した[10]。セレノによる2005年の定義では、ティラノサウルス科はアルバートサウルス、ゴルゴサウルス、ティラノサウルスを含む最も包括的でない系統群として定義された[37]。
ティラノサウルス科系統の分岐分析では、よくタルボサウルスはティラノサウルスの姉妹群に置かれ、ダスプレトサウルスはそれらよりも基盤的な位置に置かれる。タルボサウルスとティラノサウルスの近縁性は多数の頭蓋骨の特徴に支持されており、特定の骨の縫合線のパターン、それぞれの目の後ろの後眼窩骨の三日月型をしたクレストの存在、下端が下側に顕著に湾曲する非常に深い上顎骨などがある[10][15]。2003年にフィリップ・J・カリーらが提唱した別の仮説では、鼻骨と涙骨を接続する骨質の先端部が存在しないことに基づき、ダスプレトサウルスはアジア産のタルボサウルスやアリオラムスを含む系統群の基盤的メンバーであるという弱い支持が示された[17]。
関連する研究でも、2属の間で共有される下顎の固定機序が指摘されている[38]。別の論文でカリーはアリオラムスがタルボサウルスの幼体である可能性を指摘したが、より卓越したアリオラムスの鼻骨クレストと本数の多い歯から別属であることが示唆されているとも主張した。同様に、カリーはナノティラヌスの歯の本数を用いて同属がティラノサウルスと別属である可能性を示唆した[11]。しかし、キアンゾウサウルスの発見・記載により、アリオラムスはタルボサウルスと近縁ではなく、新たに記載されたアリオラムス族に属するとされた。さらに、キアンゾウサウルスの存在からは、吻部の長いティラノサウルス科の恐竜がアジアに幅広く分布しており、また異なる獲物を狩ることでより大型かつ頑強なティラノサウルス亜科との競合を回避しつつ環境を共有していたことが明らかになった[39]。
|
|
|
古生物学
成長

古生物学者グレゴリー・エリクソンらはティラノサウルス科の成長と生活史を研究した。ボーンヒストロジーの分析からは標本が死亡した際の年齢を決定でき、様々な個体の年齢を彼らの大きさに対してグラフ上にプロットすれば成長率を調査できる。エリクソンは、ティラノサウルスが長い幼体期間を過ごした後、一生涯の途中の約4年間に驚異的な成長期を迎えたことを明らかにした。性成熟とともに急成長期が終わると、成体では成長がかなり鈍化する。ティラノサウルス類の成長曲線はS字型であり、14歳前後での個体の成長速度が最大となる[43]。

既知のティラノサウルス・レックスの個体で最小のものは LACM 28471であるが、これは体重僅か29.9キログラムの2歳の個体である。この一方、最大級の個体の1つとされているものが FMNH PR2081(スー)であり、体重は約5654キログラム、年齢は28歳と推定されており、本種の上限に近かった可能性がある[43]。ティラノサウルス・レックスの幼体は約14歳になるまで体重1800キログラム未満であり、約14歳で劇的に体サイズを増大する。この急速な成長期の間に、若いティラノサウルス・レックスは4年間で年間平均600キログラムの体重増加を経験する。この成長速度は16歳から低下し、18歳で再びプラトーに達して劇的に成長が減速する[44]。例えば、28歳のスーと22歳のカナダの標本(RTMP 81.12.1)の間では僅か600キログラムの差異しかない[43]。この急激な成長速度変化は肉体的な成熟を示唆する可能性がある。この仮説はモンタナ州産の18歳のティラノサウルス・レックスの標本(MOR 1125、Bレックス)の大腿骨から骨髄組織が発見されたことで支持されている[45]。骨髄組織は排卵期の雌の鳥類でのみ確認されており、このことからBレックスは繁殖期にあったことが示唆される[46]。
他のティラノサウルスも酷似した成長曲線を示すが、成体のサイズがティラノサウルスと比較して小さいため、成長速度もそれに対応する[47]。アルバートサウルス亜科と比較してダスプレトサウルスは急速成長期における成長速度が高いが、これは成体の体重がより重いためである。成体体重1800キログラムという推定に基づくと、ダスプレトサウルスの最大成長速度は年間180キログラムである。他の研究者はダスプレトサウルスの体重をより重く見積もっているため成長速度の規模は左右されうるが、全体のパターンに変化はないと考えられる[43]。最も若い既知のアルバートサウルスはドライアイランドのボーンベッドで発見された2歳の個体で、体重は約50キログラム、と推定され、全長は2メートルを僅かに上回る。同じ産地からは既知で最大かつ最高齢の個体が発見されており、全長は10メートル、年齢は28歳である。最速成長速度は12 - 16歳で迎えたと推定され、体重1300キログラムの成体に基づき、その速度は年間122キログラムに達したと見積られる。これはティラノサウルス・レックスの成長速度の五分の一である。ゴルゴサウルスでは、計算された最大成長速度は急成長段階において約110キログラムであり、アルバートサウルスのものに匹敵する[43]。
いまだ未確認の属のティラノサウルス類の胚が発見されており、ティラノサウルス科は卵の中での発生段階で特徴的な骨格構造を発達させることが示唆される。さらに、1983年にモンタナ州のツーメディスン層で発見された下顎の1.1インチ(2.8cm)の歯と、2018年にホースシューキャニオン層で発見され2020年に記載された足の鉤爪の標本の大きさから、ティラノサウルス科の新生児はマウスやそれに近い大きさの齧歯類の全身と同等規模の頭蓋骨を伴って生まれ、全身はほぼ小型犬程度の大きさだった可能性があると考えられる。下顎の標本は全長約0.76メートルの個体、鉤爪は全長約0.91メートルの個体のものであると考えられている。いずれの標本も関連する卵殻は未発見であるが、これらのティラノサウルス科の新生児が発見された場所はこの動物が共存・捕食した他の種と同じ巣を利用していたことが示唆される[48]。また、卵殻がこれらの標本に無いことから、ティラノサウルス科はムササウルス属やプロトケラトプス属がそう考えられているように、軟殻の卵を産んだ可能性が指摘されている[49]。
ワピチ層の足跡化石からは、ティラノサウルス科は爪先が厚くなって足が幅広になり、体重を支持するように成長したことが示唆される。幅広の足は、成体のティラノサウルス科が子孫よりも緩慢に動いていたことを示唆する[50][51]。
生活史

急成長段階の終わりからは、アルバートサウルスは性成熟期に入ったことが示唆される。ただし、その後も一生涯を通じて緩慢に成長を続けたことも示唆される[43][47]。まだ活発に成長している時期に性的に成熟することは小型[52]あるいは大型[53]恐竜だけでなく、ヒトやゾウといった大型哺乳類にも共通する形質のようである[53]。相対的に性成熟の早いパターンは、成長が完了した後まで性成熟を遅延させる鳥類のパターンとは顕著に異なる[53][54]。
各年齢層の標本数を集計することにより、エリクソンらはティラノサウルス科の個体群における生活史について結論を導いた。その結果、化石の記録では幼体は稀である一方、急成長期にある亜成体や成体は遥かに多いことが判明した。半数を上回るティラノサウルス・レックスの標本は性成熟後6年以内に死亡しているようであり、これは他のティラノサウルス類や、現代の大型かつ長寿の鳥類や哺乳類にも見られるパターンである。これらの種は新生児の時期の死亡率が高く、次いで幼体の時期の死亡率が比較的低いことが特徴である。性成熟後には死亡率が再び上昇するが、これは生殖のストレスが一因である。これは保存状態や採集のバイアスに起因する可能性もあるが、エリクソンは、ゾウなど現代の大型哺乳類にも見られる一定以上の大きさの幼体の死亡率が低いことが原因ではないかと推論した。ティラノサウルスは2歳までに同時代の全ての捕食者を上回る大きさになるため、捕食者の不在に起因して死亡率が低下した可能性がある。古生物学者が十分な数のダスプレトサウルスの化石を発見していないため同様の分析は不可能であるが、エリクソンは同様の一般的な傾向があるようだと指摘した[47]。
ティラノサウルス科はその生涯の半分を幼体として過ごした後、わずか数年でほぼ最大サイズまで急成長を遂げる[43]。このことと、巨大な成体のティラノサウルス科と他の小型獣脚類の中間的な大きさの捕食者が完全に欠落していることから、これらの生態的地位は幼体のティラノサウルス科が埋めていたことが示唆される。これは孵化した後に樹上生活を送り、緩慢に成熟して大型脊椎動物を引き倒すことの出来る頑丈な頂点捕食者に至る、現生のコモドオオトカゲにも見られる[10]。例えば、アルバートサウルスは群集で発見されているが、その群れには異なる年齢の個体が入り混じっている[55][56]。
移動
移動能力が最もよく研究されているのはティラノサウルスについてであり、これに関して2つの話題がある。1つはどれほど旋回が得意であったか、もう1つは直線運動の最高速度はどれほどであったか、である。ティラノサウルスの旋回は緩慢であった可能性があり、45度回転するのに1~2秒を要したと考えられる。45度とは、直立していてかつ尾のない人間が1秒で回転可能な量である[57]。旋回を苦手とする原因は回転慣性にある。これは、ティラノサウルスの質量の多くは、人間が重い材木を運ぶように、重心からある程度離れたところに存在したためである[58]。
研究者による最大速度の推定は非常に値の幅が広く、主に約11メートル毎秒と推定されているが、5 - 11メートル毎秒や20メートル毎秒とする推定値もある。この原因は、歩行した痕跡は数多く発見されている一方で走行した痕跡は発見されておらず、彼らが走行しなかったことを示す可能性があるからである[59]。

1993年、ジャック・ホーナーとドン・セッレムはティラノサウルスが緩慢でありおそらく走行できなかった(脚が空中に浮かぶフェーズが存在しない)と主張した[60]。しかし、Holtz (1998) はティラノサウルス科およびその近縁な動物は最速の大型獣脚類だったと結論した[61]。Christiansen (1998) はティラノサウルスの脚の骨がトップスピードが限定されていてかつ実際に走行できないゾウのものよりも遥かに強靭であるわけではないと推定し、ティラノサウルスの最高速度を約11メートル毎秒と提唱した。これはヒトのスプリンターの速度と同程度である[62]。Farlow ほか (1995) は6 - 8トンに達するティラノサウルスが高速運動中に転倒すれば、地面に叩き付けられた際に小さな前肢では衝撃を減衰させられず重症あるいは致命傷を負ったと主張した[63][64]。しかし、動物園のような安全な環境に置かれていたとしても、キリンは骨折の危険がありながらも50キロメートル毎時でギャロップを行うことが知られている[65][66]。このため、ティラノサウルスも必要に応じて高速で動き、そのような危険を受け入れていた可能性もある。このシナリオはアロサウルスにおいても研究されている[67][68]。さらに後の研究では、ティラノサウルスの走行速度は17キロメートル毎時から40キロメートル毎時、すなわち歩行や緩慢な走行から中速の走行までで、それ以上に絞り込まれてはいない[59][69][70]。化石から直接得られたデータを用い2007年にコンピュータモデル研究によって推定された走行速度は8メートル毎秒であった[71][72]。
Eric Snively らが2019年に発表した研究では、タルボサウルスやティラノサウルスといったティラノサウルス科は同サイズのアロサウルス上科よりも機動性が高いことが示唆された。これは、大型の後肢の筋肉に結び付く体重と比較して回転慣性が小さいためである。結果として、ティラノサウルス科は相対的に素早い旋回が可能で、獲物に近づく際あるいは他に旋回する際にはそういった動作を見せた可能性が高く、または旋回中に片脚を固定しながらもう片方を持ち上げる動作をした可能性があるとの仮説が立てられた。この研究結果は、ティラノサウルス類の進化に敏捷性が寄与した過程を明らかにする可能性がある[73]。
加えて、2020年の研究ではティラノサウルス科は例外的に効率的な歩行者であったことが示唆された。Dececchi et al. によると、ティラノサウルス科を含む70種を超える獣脚類恐竜の後肢・体重・足取りが比較された。調査チームは様々な方法を適用してそれぞれの恐竜の最高速度、および歩行時のように緩やかな速度での運動時に消費したエネルギー量を推定した。ドロマエオサウルス科などより小型から中型の種の間では、より長い後肢はより速い走行への適応のように見え、他の研究者による以前の結果と整合する。しかし、体重1000キログラムを超える獣脚類では、最高走行速度は体サイズによる制限を受けており、そのためより長い後肢は速度ではなく低エネルギーの歩行に相関することが判明した。さらに研究結果からは、より小型の獣脚類は進化した長い脚を狩りと捕食者からの逃走の両方に役立てていた一方、大型の捕食性獣脚類は頂点捕食者としての役割ゆえに捕食圧から解放されており、進化した長い脚をエネルギーコストの減少と採餌の効率に役立てていたことが示唆された。研究でより基盤的な獣脚類のグループと比較されたティラノサウルス科は、狩りと死肉漁りの間のエネルギー支出が減少したため、採餌効率の顕著な上昇を示した。これは結果としてティラノサウルス類が彼ら自身を維持するために必要な狩りの襲撃と食餌の量を低減することに帰結した可能性が高い。加えて彼らの研究では、ティラノサウルス類が他の大型獣脚類よりもより俊敏であったことを示す研究との繋がりの中で、長距離に亘って獲物へ忍び寄って接近した後に突発的な急スピードで以て殺害することにティラノサウルス類が長けていることが示唆されている。結果としてティラノサウルス科と現生のオオカミの間に類似性が指摘されており、これはアルバートサウルスなどの少なくとも一部のティラノサウルス科が集団で狩りを行っていたことによって裏付けられている[74][75]。
外皮

古生物学コミュニティで続いている議論として、ティラノサウルス科の外皮の被覆の範囲と特性に関するものがある。前期白亜紀にあたる義県累層や中国の遼寧省に分布するその他の層からは、無数のコエルロサウルス類の骨格と共に長いタンパク質繊維が保存され、産出している[76]。他の仮説も提唱されているものの[77]、通常これらの繊維は鳥類および非鳥類型獣脚類に見られる枝分かれした羽毛とのホモログであるプロトフェザーとして解釈されている[78][79]。2004年に記載されたディロングの骨格はプロトフェザーを含んでおり、これはティラノサウルス上科において確認された最初の例である。現生鳥類のダウンフェザーと同様にディロングに見られたプロトフェザーは枝分かれしていたものの、大羽 (en) にはなっておらず、体温の保温に用いられていた可能性がある[20]。2012年には全長9メートルに達するユウティラヌスが記載され、本属の発見からは大型獣脚類の成体も羽毛が生えていた可能性が示唆された[80]。

系統ブラケッティングの原則に基づき、ティラノサウルス科はそのような羽毛を持った可能性があると予想された。しかし2017年の研究では、アルバータ州・モンタナ州およびモンゴル国で収集された5属(ティラノサウルス、アルバートサウルス、ゴルゴサウルス、ダスプレトサウルス、タルボサウルス)のティラノサウルス科の皮膚の印象化石が記載された。皮膚の印象化石は小型であるものの、これらは頭部よりも後側に幅広く分布しており、腹部・胸部・腸骨・骨盤・尾部・頸部に位置していた。これらは重複していない細かく小石様の密な鱗を示しており、羽毛の痕跡は無かった。基本的なテクスチャーは直径約1 - 2mmの小さな "basement scales" で構成され、その間に約7mmの "feature scales" の印象が散見される[81]。ティラノサウルス類の足跡にもさらなる鱗が見られる[82]。ティラノサウルス類の顔面の外皮は歯骨と上顎骨に鱗があり、表皮は角質化し、下位領域には鎧のような皮膚があることが研究で判明している[83][84]。
Bell et al. はティラノサウルス上科の外皮の分布について既知の内容に基づいて祖先形質復元を行った。ティラノサウルス上科の起源に羽毛があることの信頼性は89%であったにも拘わらず、ティラノサウルス科が鱗を持つ信頼性は97%で真と判断された。研究チームはこのデータが「ティラノサウルスが完全に扁平上皮で覆われていたことの説得力ある証拠をもたらす」とした。ただし、皮膚の印象が未発見である背中側にはまだ羽毛が存在していた可能性があると彼らは認めている。Bell et al. は、裏付ける根拠を必要としつつも、ティラノサウルス類の鱗の印象が羽毛から二次的に派生したものである可能性があるという仮説を立てている[81]。しかし、ティラノサウルス科におけるタフォノミーバイアスに起因するという主張もなされている[85]。
そのような外皮が何故発生したかはまだ決定されていない。羽毛が喪失した前例は鳥盤類などの他の恐竜のグループにも見られている。その過程においては、繊維状構造が失われ、鱗が再出現したのである[86]。この機序としては大型化が提唱されているが、フィル・R・ベルは羽毛のあるユウティラヌスとゴルゴサウルスおよびアルバートサウルスの体サイズが近いことを指摘した。ベルは体サイズと気候が類似することを挙げ、差異の理由が分からないとコメントした[87]。
視界

ティラノサウルスは眼窩が前側に着くことから、その眼球は前側を向き、現生のタカよりも僅かに良い両眼視機能を持った。一般的な捕食性獣脚類は頭蓋骨の真正面に両眼視機能を持つが、ティラノサウルスは広範囲で視野が重複した。ジャック・ホーナーは、ティラノサウルス類の系統が着実に両眼視機能を向上させてきたことを指摘した。ティラノサウルス類が純粋な腐肉食動物(スカベンジャー)であった場合、立体視をもたらす発達した奥行感覚は不要であり、この長期的な傾向が自然選択に好まれることは考えにくい[88][89]。現生の動物では、立体視は主に捕食動物に見られる。ティラノサルスと異なり、タルボサウルスは他のティラノサウルス科に典型的なより狭い頭蓋骨を持ち、眼球は主に横に面していた[90]。ゴルゴサウルスの標本では、眼窩は楕円形あるいは鍵穴型というよりもむしろ円形であり、他のティラノサウルス科の属と異なる[11]。ダスプレトサウルスでは、これは上下に高い楕円形であり、ゴルゴサウルスに見られる円形とティラノサウルスに見られる鍵穴型の中間である[10][11][91]。
顔面の感覚
ダスプレトサウルスと現生のワニのボーンテクスチャの比較に基づき、トーマス・D・カールらの2017年の詳細な研究でティラノサウルス科は吻部に大型で平坦な鱗を持つことが判明した[92][93]。鱗の中心にはケラチン質の小型のパッチが存在した。ワニにおいては、そのようなパッチは化学的刺激・熱的刺激・機械的刺激を検知可能な感覚ニューロンの束を被覆する[94][95]。彼らは、ティラノサウルス科がおそらく顔面の鱗の下に感覚ニューロンの束を有しており、ティラノサウルス科はそれを用いて対象を特定し、巣の温度を計測し、静かに卵や幼体を持ち上げた可能性があると提唱した[92]。
骨質のクレスト

数多くのティラノサウルス科恐竜を含め、多数の獣脚類の頭蓋骨には骨質のクレストが見られる。モンゴルから産出したアリオラムスは、鼻骨上に卓越した5個の骨質の隆起からなる1本の列を持つ。より隆起が低いものの、同様の列はアパタチオサウルスの頭蓋骨にも存在し、ダスプレトサウルスやアルバートサウルスおよびタルボサウルスの一部の標本にも見られる[15]。アルバートサウルスとゴルゴサウルスおよびダスプレトサウルスにおいては、涙骨上の眼の正面に卓越した角が存在する。涙骨の角はタルボサウルスとティラノサウルスには存在せず、その代わりに後眼窩骨上の眼の後方に三日月形のクレストが存在する。これらの頭部のクレストはディプレイに用いられた可能性があり、種の認識や求愛行動に用いられた可能性もある[10]。
体温調節
大部分の恐竜と同様にティラノサウルスは長らく外温性と考えられていたが、1960年代後半から始まった恐竜ルネサンスの初期にロバート・T・バッカーおよびジョン・オストロムなどの研究者はこの見解に対し異議を唱えた[96][97]。ティラノサウルス・レックスは内温性であるとされ、非常に活発な生態が示唆される[98]。それ以降、複数の古生物学者がティラノサウルスの体温調節能力を明らかにしようと努めてきた。若いティラノサウルス・レックスが哺乳類や鳥類と比較して高い成長率を持っていたヒストロジー的証拠は、高い代謝を有した仮説を示唆する可能性がある。哺乳類や鳥類におけるものと同様に、ティラノサウルス・レックスの成長曲線からは、本種の成長が主として未成熟個体に制限されていたことが示唆される。これは永続的に成長を遂げる他の脊椎動物の大半とは異なる[44]。胴椎と脛骨の体温差は4 - 5℃を以下であったことが示唆される。体の中枢と末梢での温度範囲が小さいことから、古生物学者リース・バリックと地球化学者ウィリアム・シャワーズは、ティラノサウルス・レックスの体内温度は恒常的であり、外温性の爬虫類と内温性の哺乳類の中間程度の代謝を有していたとした[99]。後に、数百万年過去の時代で別の大陸に生息していたギガノトサウルスの標本においても彼らは同様の結果を発見した[100]。もしティラノサウルス・レックスに体温を恒常的に維持した証拠が無いとしても、それは外温性であったことを必ずしも意味しない。このような体温調節は現生のウミガメのように、大型動物ゆえの保温により説明される[101][102][103]。
古生態
ダスプレトサウルスとゴルゴサウルスの共存

ダイナソーパーク累層において、ゴルゴサウルスはより希少なティラノサウルス亜科の種であるダスプレトサウルスと共存した。これは2属のティラノサウルス類が共存した数少ない例である。現生の捕食動物のギルドにおいて、同様の体サイズの捕食動物は競争を制限する解剖学的・行動学的・地理学的な差異により異なる生態的地位(ニッチ)に分裂する。ダイナソーパーク累層のティラノサウルス科におけるニッチ分割は多くが不明である[104]。1970年、デイル・ラッセルは、より個体数が少なく狩りが困難である角竜類と曲竜類がダスプレトサウルスに残されていたと仮説を立てた[12]。しかし、トゥーメディスン累層から産出したダスプレトサウルスの標本 OTM 200 には、その腹部に消化されたハドロサウルス類の幼体の遺骸が保存されている[105]。他のいくつかの獣脚類のグループと異なり、どちらの属も標高差による化石産出量の多寡は認められない[104]。しかし、ゴルゴサウルスはダイナソーパーク累層のような北方の層でより一般的であり、ダスプレトサウルスの種は南でより豊富である。同様のパターンは他の恐竜のグループにも見られる(カンパニアン期の北アメリカ南西のカスモサウルス亜科およびハドロサウルス亜科、北アメリカ北部のセントロサウルス亜科およびランベオサウルス亜科)。ホルツは、このパターンはティラノサウルス亜科・カスモサウルス亜科・ハドロサウルス亜科の間で生態的嗜好性が共有されていることを示唆しているとした。マーストリヒチアン期の末には、ティラノサウルス・レックスのようなティラノサウルス亜科、エドモントサウルスのようなハドロサウルス亜科、トリケラトプスのようなカスモサウルス亜科が北アメリカ西部に広く分布した。その一方、アルバートサウルス亜科とセントロサウルス亜科は絶滅し、ランベオサウルス亜科は衰退した[10]。
社会的行動
ティラノサウルス科の間で社会的行動があった証拠も僅かに存在する。スーの標本と同じ産地では亜成体および幼体の骨格が発見・報告されており、ティラノサウルス類がある種の社会集団の中で生活していたという仮説の支持に用いられている[106]。ゴルゴサウルスには群居性の行動の証拠は存在しないが[55][56]、アルバートサウルスやダスプレトサウルスは群れで行動した証拠が存在する[107]。
ダイナソーパーク累層のダスプレトサウルスの種の若い標本 TMP 94.143.1 では、他のティラノサウルス類に起因する噛み後が顔に存在する。噛み跡は治癒しており、噛まれた後も個体が生き延びたことが示唆される。完全に成長したダイナソーパーク累層のダスプレトサウルス TMP 85.62.1 もティラノサウルス類の噛み跡を示しており、顔面への攻撃が若い個体のみに限ったものではないことが示唆される。噛み後は他の種のものである可能性もあるものの、顔面への噛み付きなど種内攻撃は捕食動物の間で非常によく見られることである。顔面への噛み付きはゴルゴサウルスやティラノサウルスといった他のティラノサウルス類だけでなく、シンラプトルやサウロルニトレステスといった他の獣脚類の属にも見られる。ダレン・ダンケとフィリップ・カリーは、噛み跡は縄張りや資源、あるいは社会集団内の支配をを巡る種内競争に起因すると仮説を立てた[55]。

ダスプレトサウルスが社会集団として生息していた証拠は、モンタナ州のトゥーメディスン累層のボーンベッドから得られている。ボーンベッドには3頭のダスプレトサウルスの遺骸を含んでおり、これらは大型の成体・小型の幼体・中間サイズの個体からなる。同産地には少なくとも5頭のハドロサウルス類が保存されている。地質学的証拠からは、遺骸は川の流れによって集積されたのではなく、全ての動物が同じ場所に同時に埋没したことが判明している。ハドロサウルス類の遺骸は分散しており、またティラノサウルス類の歯による多数の噛み跡が残されている。これは死亡時にダスプレトサウルスがハドロサウルス類を採餌していたことを示唆している。カリーは、確証を持つことはできないもののダスプレトサウルスが群れを形成していたことを推測した[56]。他の研究者は、ダスプレトサウルスや他の大型獣脚類が社会集団を形成した証拠について懐疑的である[108]。ブライアン・ローチとダニエル・ブリンクマンは、ダスプレトサウルスの社会相互作用は現生のコモドオオトカゲのものに類似するものであるとした。コモドオオトカゲの群れは非協力的であり、死骸にありつき、その過程で互いに共食いすることも頻繁にある[109]。
バーナム・ブラウンらが発見したドライアイランドのボーンベッドには、22頭のアルバートサウルスの遺骸が保存されている。これは白亜紀の獣脚類の中で最も多くの個体が1つの場所で発見された例であり、大型獣脚類の中ではユタ州のクリーブランド=ロイド恐竜採石場のアロサウルス群集に次いで2番目に多い。この集団には非常に高齢な1頭の成体、17歳から23歳の8頭の成体、12歳から16歳で急成長段階にある7頭の亜成体、2歳から11歳の成長段階に達していない6頭の幼体が含まれる[47]。植物食性動物の遺骸がほぼ存在しないこと、およびアルバートサウルスのボーンベッドでの数多くの個体の保存状態との類似から、フィリップ・カリーはこの産地がカリフォルニア州のラ・ブレア・タールピットのような捕食動物の罠だったのではなく、保存された全ての動物が同時に死亡したと結論した。カリーはこれを群れとしての行動の根拠とした[110]。他の研究者は懐疑的であり、旱魃や洪水あるいは他の要因により動物たちが運搬されたと解釈した[47][108][111]。
一般に議論が続いているものの、少なくともいくつかのティラノサウルス科の種が社会的であったという仮説を支持する証拠は存在する。ブリティッシュコロンビア州のワピチ累層では、地元住民のアーロン・フレッドランドにより3頭のティラノサウルス科の個体に由来する足跡が発見され、リチャード・マッケラらにより記載された。足跡の調査からは、1つの足跡が作られた後に長期間放置された痕跡が認められず、3頭のティラノサウルス類の個体が集団で移動していたという仮説が支持された。さらなる調査では、これらの動物が時速6.3 - 8.4キロメートルで移動し、腰の高さは2.1 - 2.7メートルであった可能性が高いことが判明した。この層からはティラノサウルス科の異なる3属(ゴルゴサウルスとダスプレトサウルスおよびアルバートサウルス)が知られており、どの属が足痕を残したかは不明である[112][113][114]。さらなる証拠は、ユタ州南部のカイパロウィッツ累層のRainbows and Unicorns Quarryのボーンベッドにある。この化石証拠はテラトフォネウスのものとされており、2021年に記載され、他のティラノサウルス科も社会的動物であったことが示唆された。4 - 22歳に亘る異なる4 - 5頭の動物からなるこの化石からは、おそらく洪水、あるいはより低い可能性で藍藻の毒や火災や旱魃によって引き起こされた大量死事変が示唆されている。テラトフォネウスやアルバートサウルス、ティラノサウルスやダスプレトサウルスといった属のボーンベッドはティラノサウルス科に広く社会的行動が見られたことを意味しており、保存された全ての動物が短期間に死滅したという事実はティラノサウルス科の群居性行動の論拠を補強している[115][116][117]。
食餌
ティラノサウルス類の歯型は肉食恐竜の食痕として最も広く保存されており[118]、角竜類やハドロサウルス類および他のティラノサウルス類から報告されている[118]。既知のティラノサウルス科の骨のち約2%には歯型が保存されている[118]。ティラノサウルス科の歯はナイフのように切断する機能よりもむしろ肉を体から引き剥がす金具のように用いられた[119]。歯の摩耗パターンからは、ティラノサウルス類の食餌には頭を振る複雑な行動が含まれていたことが示唆される[119]。

少数の研究者は、アルバートサウルスの群れでの狩猟行動について、群れの若いメンバーが獲物をより大型かつ強力であるが緩慢な成体まで誘導したと推測した[110]。幼体は成体と異なる生態を持っていた可能性があり、巨大な成体とより小型の同時代の獣脚類との間のニッチを生めていた可能性がある。同時代の小型獣脚類は、その最大の種でも成体のアルバートサウルスよりも体重が2桁小さいのである[10]。しかし、化石記録に行動が保存されることは非常に稀であり、これらのアイディアは容易に検証が可能なものではない。フィリップ・カリーは、ダスプレトサウルスが群れを形成して狩りを行ったと思弁したが、確証を持っての主張は出来ないとした[56]。ゴルゴサウルスでそのような行動の証拠は発見されていない[55][56]。
ティラノサウルスが捕食動物であったか腐肉食動物であったかという議論は、そのロコモーションに関する議論と同程度に昔からのものである。Lambe (1917) はティラノサウルスに近縁なゴルゴサウルスの骨格を記載し、ゴルゴサウルスの歯がほぼ摩耗していないことに基づいてゴルゴサウルスおよびティラノサウルスが純粋な腐肉食動物であったと結論した[120]。獣脚類の歯は極めて急速に置換されるため、この主張は真剣には受け止められていない。ティラノサウルスの最初の発見以降大半の研究者はティラノサウルスが捕食動物であったことに同意しているが、現生の捕食動物のように機会があれば喜んで他の捕食者の獲物を盗んだり死肉を漁ったりしたと思われる[121][122]。
ジャック・ホーナーは、ティラノサウルスが専ら腐肉食動物であり、一切の狩りを行わなかったと考えた[60][123][124]。ホーナーは純粋腐肉食動物仮説を支持する複数の主張を紹介している。大型の嗅球と嗅神経の存在からは、遠距離の死骸を嗅ぎ付けるための高度に発達した嗅覚が示唆される。歯は骨を砕くことができ、それゆえ通常は最も栄養分の少ない部位である死体の残りから可能な限りの食料(骨髄)を抜き取ることが可能だった。また、ティラノサウルスは走るのではなく歩いていたことが証拠から示唆されているが、少なくともその獲物となりうる動物は俊敏に動くことが出来た[123][125]。
他の証拠からは、ティラノサウルスの狩猟行動が示唆されている。ティラノサウルス類の眼窩は目が前を向くように位置しており、良好な立体視が可能であった。ティラノサウルス類が負わせた負傷がトリケラトプスやハドロサウルス類の骨格に認められており、それらは最初の攻撃を生き延びたように見える[126][127][128]。ティラノサウルスが腐肉食動物ならば、北アメリカからユーラシアにかけての上部白亜系に別の恐竜が頂点捕食者の地位に立っていなければならないと主張する研究者もいる。頂点捕食者の獲物は大型の周飾頭類と鳥脚類であった。他のティラノサウルス科はティラノサウルスと数多くの特徴を共有するため、ティラノサウルスを除外した場合、頂点捕食者の候補として残るのは小型であるドロマエオサウルス類のみである。このことから、腐肉食動物仮説を信じる者は、ティラノサウルス類がその巨体と力で以て小型動物から獲物を盗むことが可能だったと提唱した[125]。
共食い
ティラノサウルス科が少なくとも時折共食いしていたことを強く示唆する証拠もある。ティラノサウルス自体には1つの標本において足・上腕骨・中足骨に歯型が見られるものがあり、これに基づいて少なくとも死骸を漁るために共食いが行われていたことが示唆される[129]。フルートランド層およびキルトランド層(共にカンパニアン期)産の化石、およびオジョ・アラモ層(マーストリヒチアン期)産の化石からは、San Juan 盆地の様々なティラノサウルス科で共食いが起きていたことが示唆される。標本から集められた証拠に基づくと、ティラノサウルス科が同種の動物の共食いを伴う日和見的な摂食行動をとっていた[130]。
分布

初期のティラノサウルス上科が北半球の全ての大陸で発見されている一方、ティラノサウルス科の化石は北アメリカとアジアでしか発見されていない。南半球で産出した断片的な化石がティラノサウルス科のものとして報告されることは複数回あったが、これらはアベリサウルス科の化石の誤同定である[131]。白亜紀の中頃の化石記録がアジア・北アメリカの両地域で乏しいためティラノサウルス科の起源となった時代と場所は不明なままであるが、ティラノサウルス科のものとして確認された最初期のものはカンパニアン期の北アメリカ西部に生息していた[10]。
北アメリカ東部でティラノサウルス科の化石は発見されていないが、ドリプトサウルスやアパラチオサウルスといったより基盤的なティラノサウルス上科の化石は回収されている。これらは白亜紀の終わりまで持続しており、このことから白亜紀の中頃に西部内陸海路によって北アメリカが分裂した後、西部でティラノサウルス科が進化したことが示唆される[15]。ティラノサウルス科の化石はアラスカ州からも発見されており、これが北アメリカとアジアに分散したルートであった可能性がある[132]。ある系統解析ではアリオラムスとタルボサウルスは近縁な位置に置かれており、本科において固有のアジアの系統を形成した[17]。後にキアンゾウサウルスの発見およびアリオラムス族の記載によりこれは反証された[29]。2015年、日本の福井県立恐竜博物館は長崎県長崎市の長崎半島から産出したティラノサウルス科の歯化石を報告した。これは8100万年前(カンパニアン期)のもので、日本国内初の大型ティラノサウルス類の歯は化石として発表された[133]。

ティラノサウルス亜科はより広範囲に分布した。アルバートサウルス亜科はアジアで発見されておらず、アジアにはタルボサウルスやズケンティラヌスおよびアリオラムス族(アリオラムスやキアンゾウサウルス)といったティラノサウルス亜科が生息していた。ティラノサウルス亜科とアルバートサウルス亜科は共にカンパニアン期 - マーストリヒチアン期に北アメリカに生息しており、ダスプレトサウルスのようなティラノサウルス亜科は西部内陸中に分布した一方、アルバートサウルス亜科のアルバートサウルスやゴルゴサウルスは大陸の北西部のみでしか知られていない[134]。
後期マーストリヒチアン期までにアルバートサウルス亜科は絶滅し、ティラノサウルス亜科がサスカチュワン州からテキサス州まで分布した。このパターンは他の北アメリカの恐竜にも当てはまっている。カンパニアン期から前期マーストリヒチアン期にかけて、北西部ではランベオサウルス亜科の鳥脚類とセントロサウルス亜科の角竜が繁栄し、南部ではハドロサウルス亜科とカスモサウルス亜科が繁栄した。白亜紀末ではセントロサウルス亜科は発見されておらず、ランベオサウルス亜科も数を減らしていた一方、ハドロサウルス亜科とカスモサウルス亜科は西部内陸中で繁栄した[10]。2016年にスティーヴ・ブルサッテらは、北アメリカ西部の大部分の他のティラノサウルス科の絶滅にティラノサウルス自体が部分的に関与していたことを示唆した。この研究では、ティラノサウルスが北アメリカで進化したのではなく(おそらくタルボサウルスの子孫として)アジアから渡り、他のティラノサウルス科と競合して生存競争に勝利したことが示唆されている。この仮説は、ティラノサウルスが知られている範囲で他種のティラノサウルス科がほとんど発見されていない事実からも支持されている[135]。
属の時系列
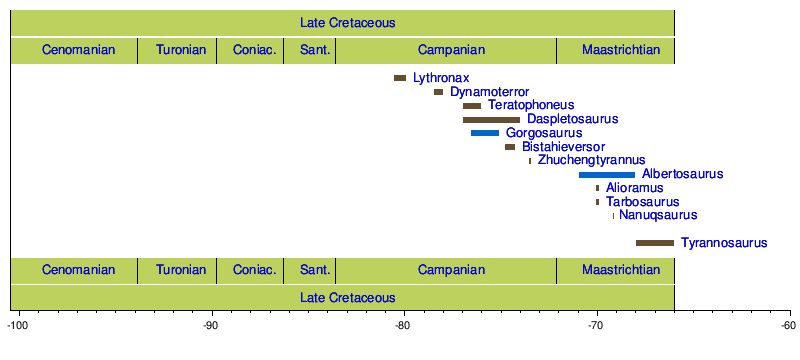
出典
- ^ a b グレゴリー・ポール 著、東洋一、今井拓哉、河部壮一郎、柴田正輝、関谷透、服部創紀 訳『グレゴリー・ポール恐竜事典 原著第2版』2020年8月30日、116-117頁。ISBN 978-4-320-04738-9。
- ^ a b c Hutchinson, John R.; Bates, Karl T.; Molnar, Julia; Allen, Vivian; Makovicky, Peter J.; Claessens, Leon (2011). “A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth”. PLOS ONE 6 (10): e26037. Bibcode: 2011PLoSO...626037H. doi:10.1371/journal.pone.0026037. PMC 3192160. PMID 22022500.
- ^ Therrien, F.; Henderson, D. M. (2007). “My theropod is bigger than yours ... or not: estimating body size from skull length in theropods”. Journal of Vertebrate Paleontology 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2. ISSN 0272-4634.
- ^ Hartman, Scott (July 7, 2013). “Mass estimates: North vs South redux”. Scott Hartman's Skeletal Drawing.com. October 12, 2013時点のオリジナルよりアーカイブ。August 24, 2013閲覧。
- ^ a b c d e Carpenter, Ken. (1992). “Tyrannosaurids (Dinosauria) of Asia and North America”. In Mateer, Niall J.; Chen Peiji. Aspects of Nonmarine Cretaceous Geology. Beijing: China Ocean Press. pp. 250–268
- ^ Breithaupt, B.H.; Southwell, E.H.; Matthews, N.A. (18 October 2005). "In Celebration of 100 years of Tyrannosaurus rex: Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the Earliest Discoveries of Tyrannosaurus rex in the West". Abstracts with Programs. 2005 Salt Lake City Annual Meeting. Vol. 37. Geological Society of America. p. 406. 2019年10月22日時点のオリジナルよりアーカイブ。2008年10月8日閲覧。
- ^ Breithaup, BH; Southwell EH; Matthews NA (2006). “Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West”. New Mexico Museum of Natural History and Science Bulletin 35: 257–258.
- ^ Bakker R.T; Williams M.; Currie P. (1988). “Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana”. Hunteria 1: 1–30.
- ^ a b Kurzanov, Sergei M.. “A new carnosaur from the Late Cretaceous of Nogon-Tsav, Mongolia” (ロシア語). The Joint Soviet-Mongolian Paleontological Expedition Transactions 3: 93–104.
- ^ a b c d e f g h i j k l m n o p q r Holtz, Thomas R. (2004). “Tyrannosauroidea”. In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 111–136. ISBN 978-0-520-24209-8
- ^ a b c d e f g Currie, Philip J. (2003). “Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta”. Acta Palaeontologica Polonica 48 (2): 191–226. オリジナルの2018-08-19時点におけるアーカイブ。 2010年6月12日閲覧。.
- ^ a b c d Russell, Dale A. (1970). “Tyrannosaurs from the Late Cretaceous of western Canada”. National Museum of Natural Sciences Publications in Paleontology 1: 1–34.
- ^ Maleev, Evgeny A. (1955). “New carnivorous dinosaurs from the Upper Cretaceous of Mongolia” (ロシア語). Doklady Akademii Nauk SSSR 104 (5): 779–783. オリジナルの2012-11-03時点におけるアーカイブ。 2008年9月10日閲覧。.
- ^ Currie, Philip J. (2000). “Theropods from the Cretaceous of Mongolia”. The Age of Dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press. pp. 434–455. ISBN 978-0-521-54582-2
- ^ a b c d e f Carr, Thomas D.; Williamson, Thomas E.; Schwimmer, David R. (2005). “A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama”. Journal of Vertebrate Paleontology 25 (1): 119–143. doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2. ISSN 0272-4634.
- ^ Carr, Thomas D.; Varricchio, David J.; Sedlmayr, Jayc C.; Roberts, Eric M.; Moore, Jason R. (2017-03-30). “A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system” (英語). Scientific Reports 7 (1): 44942. Bibcode: 2017NatSR...744942C. doi:10.1038/srep44942. ISSN 2045-2322. PMC 5372470. PMID 28358353.
- ^ a b c d Currie, Philip J.; Hurum, Jørn H; Sabath, Karol. (2003). “Skull structure and evolution in tyrannosaurid phylogeny”. Acta Palaeontologica Polonica 48 (2): 227–234. オリジナルの2008-10-31時点におけるアーカイブ。 2010年10月2日閲覧。.
- ^ a b c d e f g Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84–89. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ^ Quinlan, Elizabeth D.; Derstler, Kraig; Miller, Mercedes M. (2007). “Anatomy and function of digit III of the Tyrannosaurus rex manus”. Geological Society of America Annual Meeting — Abstracts with Programs: 77. オリジナルの2008-02-24時点におけるアーカイブ。 2008年1月15日閲覧。. [abstract only]
- ^ a b c Xu Xing, X; Norell, Mark A.; Kuang Xuewen; Wang Xiaolin; Zhao Qi; Jia Chengkai. (2004). “Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids”. Nature 431 (7009): 680–684. Bibcode: 2004Natur.431..680X. doi:10.1038/nature02855. PMID 15470426.
- ^ Holtz, Thomas R. (1994). “The phylogenetic position of the Tyrannosauridae: implications for theropod systematics”. Journal of Paleontology 68 (5): 1100–1117. doi:10.1017/S0022336000026706. JSTOR 1306180.
- ^ Cope E.D. (1866). “Discovery of a gigantic dinosaur in the Cretaceous of New Jersey”. Proceedings of the Academy of Natural Sciences of Philadelphia 18: 275–279.
- ^ Maleev E.A. (1955). “New carnivorous dinosaurs from the Upper Cretaceous Mongolia”. Doklady Akademii Nauk SSSR 104 (5): 779–783.
- ^ Leidy, Joseph (1856). “Notice of remains of extinct reptiles and fishes, discovered by Dr. F.V. Hayden in the badlands of the Judith River, Nebraska Territory”. Proceedings of the Academy of Natural Sciences of Philadelphia 8: 72–73.
- ^ Osborn, Henry F. (1905). “Tyrannosaurus and other Cretaceous carnivorous dinosaurs”. Bulletin of the American Museum of Natural History 21 (3): 259–265. doi:10.1111/j.1468-5965.2007.00735_17.x. hdl:2246/1464.
- ^ Liddell, Henry G.; Scott, Robert (1980). Greek-English Lexicon (Abridged ed.). Oxford: Oxford University Press. ISBN 978-0-19-910207-5
- ^ Fiorillo, A. R.; Tykoski, R. S. (2014). Dodson, Peter. ed. “A Diminutive New Tyrannosaur from the Top of the World”. PLoS ONE 9 (3): e91287. Bibcode: 2014PLoSO...991287F. doi:10.1371/journal.pone.0091287. PMC 3951350. PMID 24621577.
- ^ Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 464pp. ISBN 978-0-671-61946-6
- ^ a b Lü, Junchang; Yi, Laiping; Brusatte, Stephen L.; Yang, Ling; Li, Hua; Chen, Liu (2014). “A new clade of Asian Late Cretaceous long-snouted tyrannosaurids”. Nature Communications 5: 3788. Bibcode: 2014NatCo...5.3788L. doi:10.1038/ncomms4788. PMID 24807588.
- ^ “Pinocchio rex long-snouted tyrannosaur discovered in Asia” (7 May 2014). 2 February 2019時点のオリジナルよりアーカイブ。8 May 2014閲覧。
- ^ Olshevsky, George (1995). “The origin and evolution of the tyrannosaurids”. Kyoryugaku Saizensen [Dino Frontline] 9–10: 92–119.
- ^ Olshevsky G.; Ford T.L. (1995). “The origin and evolution of the Tyrannosauridae, part 2 [in Japanese]”. Dino Frontline 6: 75–99.
- ^ Fowler, DW; Woodward, HN; Freedman, EA; Larson, PL; Horner, JR (2011). “Reanalysis of "Raptorex kriegsteini": A Juvenile Tyrannosaurid Dinosaur from Mongolia”. PLOS ONE 6 (6): e21376. Bibcode: 2011PLoSO...6E1376F. doi:10.1371/journal.pone.0021376. PMC 3126816. PMID 21738646.
- ^ Sereno, Paul C. (1998). “A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria” (ドイツ語). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210: 41–83. doi:10.1127/njgpa/210/1998/41.
- ^ a b c d e f g h Holtz, TR, Jr. (2001), The phylogeny and taxonomy of the Tyrannosauridae in K Carpenter & D Tanke [eds.], Mesozoic Vertebrate Life. Indiana Univ. Press, pp. 64-83.
- ^ Brochu, Christopher R. (2003). “Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull”. Society of Vertebrate Paleontology Memoirs 7: 1–138. doi:10.2307/3889334. JSTOR 3889334.
- ^ Sereno, Paul C. (7 November 2005). “Stem Archosauria—TaxonSearch”. 19 February 2007時点のオリジナルよりアーカイブ。14 January 2008閲覧。
- ^ Hurum, Jørn H.; Sabath, Karol. (2003). “Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared” (abstract). Acta Palaeontologica Polonica 48 (2): 161–190. オリジナルの2012-03-08時点におけるアーカイブ。 2008年9月10日閲覧。.
- ^ “Newly found dinosaur is long-nosed cousin of Tyrannosaurus rex”. ScienceDaily. 2018年3月17日時点のオリジナルよりアーカイブ。2018年3月9日閲覧。
- ^ CITEREFBrusatte_&_Carr2016
- ^ CITEREFFiorillo_&_Tykoski2014
- ^ Loewen, M.A.; Irmis, R.B.; Sertich, J.J.W.; Currie, P. J.; Sampson, S. D. (2013). Evans, David C. ed. “Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans”. PLoS ONE 8 (11): e79420. Bibcode: 2013PLoSO...879420L. doi:10.1371/journal.pone.0079420. PMC 3819173. PMID 24223179.
- ^ a b c d e f g h Erickson, Gregory M., GM; Makovicky, Peter J.; フィリップ・J・カリー; Norell, Mark A.; Yerby, Scott A.; Brochu, Christopher A. (2004). “Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs”. Nature 430 (7001): 772–775. Bibcode: 2004Natur.430..772E. doi:10.1038/nature02699. PMID 15306807. "Sampled longevities for T. rex ranged from 2 to 28 years"
- ^ a b Horner, J.R. & Padian, K. (2004). “Age and growth dynamics of Tyrannosaurus rex”. Proceedings of the Royal Society of London B 271 (1551): 1875–1880. doi:10.1098/rspb.2004.2829. PMC 1691809. PMID 15347508.
- ^ Lee, Andrew H.; Werning, Sarah (2008). “Sexual maturity in growing dinosaurs does not fit reptilian growth models”. Proceedings of the National Academy of Sciences 105 (2): 582–587. Bibcode: 2008PNAS..105..582L. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ Schweitzer, M.H., Wittmeyer, J.L., & Horner, J.R. (2005). “Gender-specific reproductive tissue in ratites and Tyrannosaurus rex”. Science 308 (5727): 1456–1460. Bibcode: 2005Sci...308.1456S. doi:10.1126/science.1112158. PMID 15933198. オリジナルの2021-11-10時点におけるアーカイブ。 2019年12月2日閲覧。.
- ^ a b c d e Erickson, G.M., Currie, P.J., Inouye, B.D., & Winn, A.A. (2006). “Tyrannosaur life tables: an example of nonavian dinosaur population biology”. Science 313 (5784): 213–217. Bibcode: 2006Sci...313..213E. doi:10.1126/science.1125721. PMID 16840697. オリジナルの2021-11-10時点におけるアーカイブ。 2019年12月2日閲覧。.
- ^ “First tyrannosaur embryo fossils revealed”. Science (October 19, 2020). December 5, 2020時点のオリジナルよりアーカイブ。November 23, 2020閲覧。
- ^ October 2020, Laura Geggel-Associate Editor 15 (15 October 2020). “1st of their kind baby tyrannosaur fossils unearthed”. livescience.com. 17 October 2020時点のオリジナルよりアーカイブ。17 October 2020閲覧。
- ^ “Fat-footed tyrannosaur parents couldn't keep up with their skinnier offspring, fossil footprints reveal”. The Conversation. 2021年4月26日時点のオリジナルよりアーカイブ。2021年4月28日閲覧。
- ^ Enriquez, Nathan J.; Campione, Nicolás E.; Brougham, Tom; Fanti, Federico; White, Matt A.; Sissons, Robin L.; Sullivan, Corwin; Vavrek, Matthew J. et al. (April 21, 2021). “Exploring possible ontogenetic trajectories in tyrannosaurids using tracks from the Wapiti Formation (upper Campanian) of Alberta, Canada”. Journal of Vertebrate Paleontology 40 (6): e1878201. doi:10.1080/02724634.2021.1878201.
- ^ Erickson, Gregory M.; Curry Rogers, Kristi; Varricchio, David J.; Norell, Mark; Xu, Xing (2007). “Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition”. Biology Letters 3 (5): 558–561. doi:10.1098/rsbl.2007.0254. PMC 2396186. PMID 17638674.
- ^ a b c Lee, Andrew H.; Werning, Sarah (2008). “Sexual maturity in growing dinosaurs does not fit reptilian growth models”. PNAS 105 (2): 582–587. Bibcode: 2008PNAS..105..582L. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ Ricklefs, Robert E. (2007). “Tyrannosaur ageing”. Biology Letters 3 (2): 214–217. doi:10.1098/rsbl.2006.0597. PMC 2375931. PMID 17284406.
- ^ a b c d Tanke, Darren H.; Currie, Philip J. (1998). “Head-biting behavior in theropod dinosaurs: paleopathological evidence”. Gaia 15: 167–184. オリジナルの2008-02-27時点におけるアーカイブ。 2008年9月13日閲覧。.
- ^ a b c d e Currie, Philip J.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; Murphy, Nate (2005). “An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)”. In Carpenter, Kenneth. The Carnivorous Dinosaurs. Bloomington: Indiana University Press. pp. 313–324. ISBN 978-0-253-34539-4
- ^ “Tyrannosaurus had poor turning circle”. May 7, 2012時点のオリジナルよりアーカイブ。2007年9月25日閲覧。
- ^ Carrier, D.R., Walter, R.M., and Lee, D.V. (15 November 2001). “Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia”. Journal of Experimental Biology 204 (Pt 22): 3917–3926. doi:10.1242/jeb.204.22.3917. PMID 11807109. オリジナルの6 January 2008時点におけるアーカイブ。 12 October 2008閲覧。.
- ^ a b Hutchinson, J.R. (2004). “Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa”. Journal of Morphology 262 (1): 441–461. doi:10.1002/jmor.10240. PMID 15352202. オリジナルの2008-10-31時点におけるアーカイブ。.
- ^ a b Horner, John R.; Don Lessem (1993). The complete T. rex. New York City: Simon & Schuster. pp. 120. ISBN 978-0-671-74185-3
- ^ Holtz, Thomas R. (1 May 1996). “Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda)”. Journal of Paleontology 70 (3): 536–538. doi:10.1017/S0022336000038506. オリジナルの26 October 2008時点におけるアーカイブ。 3 October 2008閲覧。.
- ^ Christiansen, P. (1998). “Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential”. Gaia 15: 241–255. ISSN 0871-5424. オリジナルの2008-10-31時点におけるアーカイブ。 2008年10月12日閲覧。.
- ^ Farlow, James O.; Smith, Matthew B.; Robinson, John M. (1995). “Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex”. Journal of Vertebrate Paleontology 15 (4): 713–725. doi:10.1080/02724634.1995.10011257. オリジナルの2008-10-23時点におけるアーカイブ。.
- ^ "The bigger they come, the harder they fall" Archived 2015-05-31 at the Wayback Machine. New Scientist, 7 October 1995, p. 18.
- ^ “Giraffe”. WildlifeSafari.info. 12 June 2010時点のオリジナルよりアーカイブ。29 April 2006閲覧。
- ^ “The History of Woodland Park Zoo — Chapter 4”. June 2, 2007時点のオリジナルよりアーカイブ。29 April 2006閲覧。
- ^ Alexander, R.M. (August 7, 2006). “Dinosaur biomechanics”. Proc Biol Sci 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532. PMC 1634776. PMID 16822743.
- ^ Hanna, Rebecca R. (2002). “Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd Dinosaur Quarry collection”. Journal of Vertebrate Paleontology 22 (1): 76–90. doi:10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2. ISSN 0272-4634.
- ^ Hutchinson, John R.; Garcia, M (28 February 2002). “Tyrannosaurus was not a fast runner”. Nature 415 (6875): 1018–1021. Bibcode: 2002Natur.415.1018H. doi:10.1038/4151018a. PMID 11875567.
- ^ Hajdul, R. (1997). Tendons. Dinosaur Cards. Orbis Publishing Ltd. D36044311.
- ^ Sellers, W.I. & Manning, P.L. (July 2007). “Estimating dinosaur maximum running speeds using evolutionary robotics”. Proc. R. Soc. B 274 (1626): 2711–6. doi:10.1098/rspb.2007.0846. PMC 2279215. PMID 17711833. オリジナルの2011-02-07時点におけるアーカイブ。 2008年10月12日閲覧。.
- ^ Liz Seward: "T. rex 'would outrun footballer'" Archived 2012-01-07 at the Wayback Machine.. BBC News website, Tuesday, 21 August 2007. The article quotes Dr Bill Sellers, University of Manchester, co-author of a paper published in Proceedings of the Royal Society B. Retrieved 22 August 2007.
- ^ Snively, Eric; O’Brien, Haley; Henderson, Donald M.; Mallison, Heinrich; Surring, Lara A.; Burns, Michael E.; Jr, Thomas R. Holtz; Russell, Anthony P. et al. (February 21, 2019). “Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods”. PeerJ 7: e6432. doi:10.7717/peerj.6432. PMC 6387760. PMID 30809441.
- ^ Dececchi, T. Alexander; Mloszewska, Aleksandra M.; Holtz, Thomas R.; Habib, Michael B.; Larsson, Hans C. E. (2020). “The fast and the frugal: Divergent locomotory strategies drive limb lengthening in theropod dinosaurs”. PLOS ONE 15 (5): e0223698. Bibcode: 2020PLoSO..1523698D. doi:10.1371/journal.pone.0223698. PMC 7220109. PMID 32401793.
- ^ “T. rex was a champion walker, super-efficient at lower speeds”. EurekAlert!. 2020年6月27日時点のオリジナルよりアーカイブ。2020年6月27日閲覧。
- ^ Zhou Zhonghe, Z; Barrett, Paul M.; Hilton, Jason. (2003). “An exceptionally preserved Lower Cretaceous ecosystem”. Nature 421 (6925): 807–814. Bibcode: 2003Natur.421..807Z. doi:10.1038/nature01420. PMID 12594504.
- ^ Lingham-Soliar, Theagarten; Feduccia, Alan; Wang, Xiaolin (2007). “A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres”. Proceedings of the Royal Society of London. Series B, Biological Sciences 274 (1620): 1823–1829. doi:10.1098/rspb.2007.0352. PMC 2270928. PMID 17521978.
- ^ Chen Peiji, Pei-ji; Dong Zhiming; Zhen Shuonan. (1998). “An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China”. Nature 391 (6663): 147–152. Bibcode: 1998Natur.391..147C. doi:10.1038/34356.
- ^ Xu Xing, X; Zhou Zhonghe; Prum, Richard A. (2003). “Branched integumental structures in Sinornithosaurus and the origin of feathers”. Nature 410 (6825): 200–204. Bibcode: 2001Natur.410..200X. doi:10.1038/35065589. PMID 11242078.
- ^ “Archived copy”. 2012年4月17日時点のオリジナルよりアーカイブ。2012年4月4日閲覧。
- ^ a b Bell, P. R.; Campione, N. E.; Persons, W. S.; Currie, P. J.; Larson, P. L.; Tanke, D. H.; Bakker, R. T. (2017). “Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution”. Biology Letters 13 (6): 20170092. doi:10.1098/rsbl.2017.0092. PMC 5493735. PMID 28592520.
- ^ Currie, P.J., Badamgarav, D., Koppelhus, E.B. 2003. The First Late Cretaceous Footprints from the Nemegt Locality in the Gobi of Mongolia. Ichnos. Vol.10:1-12.Currie, P.J., Badamgarav, D., Koppelhus, E.B. 2003.
- ^ Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7.
- ^ “Tyrannosaurid theropods: did they ever smile like crocodiles?”. The Annual Symposium of Vertebrate Palaeontology and Comparative Anatomy (September 2018). 2022年10月11日閲覧。
- ^ Saitta, Evan T.; Fletcher, Ian; Martin, Peter; Pittman, Michael; Kaye, Thomas G.; True, Lawrence D.; Norell, Mark A.; Abbott, Geoffrey D. et al. (2018-11-01). “Preservation of feather fibers from the Late Cretaceous dinosaur Shuvuuia deserti raises concern about immunohistochemical analyses on fossils” (英語). Organic Geochemistry 125: 142–151. doi:10.1016/j.orggeochem.2018.09.008. ISSN 0146-6380.
- ^ blogger, Guest. “Ten Things We Don't Know about Tyrannosaurs”. 2020年9月30日時点のオリジナルよりアーカイブ。2021年1月5日閲覧。
- ^ “Tyrannosaurus rex had scaly skin and wasn’t covered in feathers, a new study says”. The Washington Post (2017年6月6日). 2017年6月16日時点のオリジナルよりアーカイブ。2022年10月10日閲覧。
- ^ Stevens, Kent A. (June 2006). “Binocular vision in theropod dinosaurs”. Journal of Vertebrate Paleontology 26 (2): 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2. ISSN 0272-4634. オリジナルの2012-03-23時点におけるアーカイブ。.
- ^ Jaffe, Eric (1 July 2006). “Sight for 'Saur Eyes: T. rex vision was among nature's best”. Science News 170 (1): 3–4. doi:10.2307/4017288. JSTOR 4017288. オリジナルの29 September 2012時点におけるアーカイブ。 6 October 2008閲覧。.
- ^ Saveliev, Sergei V.; Alifanov, Vladimir R. (2005). “A new study of the brain of the predatory dinosaur Tarbosaurus bataar (Theropoda, Tyrannosauridae)”. Paleontological Journal 41 (3): 281–289. doi:10.1134/S0031030107030070.
- ^ Carr, Thomas D. (1999). “Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria)”. Journal of Vertebrate Paleontology 19 (3): 497–520. doi:10.1080/02724634.1999.10011161. オリジナルの2020-02-20時点におけるアーカイブ。 2019年12月2日閲覧。.
- ^ a b Carr, Thomas D.; Varricchio, David J.; Sedlmayr, Jayc C.; Roberts, Eric M.; Moore, Jason R. (2017-03-30). “A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system” (英語). Scientific Reports 7: 44942. Bibcode: 2017NatSR...744942C. doi:10.1038/srep44942. ISSN 2045-2322. PMC 5372470. PMID 28358353.
- ^ Barker, Chris; Naish, Darren; Katsamenis, Orestis; Dyke, Gareth (June 16, 2017). “Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii”. Scientific Reports 7 (1): 3749. Bibcode: 2017NatSR...7.3749B. doi:10.1038/s41598-017-03671-3. PMC 5473926. PMID 28623335.
- ^ Leitch, Duncan B.; Catania, Kenneth C. (2012-12-01). “Structure, innervation and response properties of integumentary sensory organs in crocodilians” (英語). Journal of Experimental Biology 215 (23): 4217–4230. doi:10.1242/jeb.076836. ISSN 0022-0949. PMC 4074209. PMID 23136155.
- ^ Di-Poï, Nicolas ; Milinkovitch, Michel C. (2013-07-02). “Crocodylians evolved scattered multi-sensory micro-organs”. EvoDevo 4 (1): 19. doi:10.1186/2041-9139-4-19. ISSN 2041-9139. PMC 3711810. PMID 23819918.
- ^ Bakker, Robert T. (1968). “The superiority of dinosaurs”. Discovery 3 (2): 11–12.
- ^ Bakker, Robert T. (1972). “Anatomical and ecological evidence of endothermy in dinosaurs”. Nature 238 (5359): 81–85. Bibcode: 1972Natur.238...81B. doi:10.1038/238081a0.
- ^ Bakker, Robert T. (1986). The Dinosaur Heresies. New York: Kensington Publishing. ISBN 978-0-688-04287-5. OCLC 13699558[要ページ番号]
- ^ Barrick, Reese E.; William J. Showers (July 1994). “Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes”. Science 265 (5169): 222–224. Bibcode: 1994Sci...265..222B. doi:10.1126/science.265.5169.222. PMID 17750663.
- ^ Barrick, Reese E.; William J. Showers (October 1999). “Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus”. Palaeontologia Electronica 2 (2). オリジナルの2011-05-17時点におけるアーカイブ。 2008年10月12日閲覧。.
- ^ Paladino, Frank V.; Spotila, James R.; Dodson, Peter (1999). “A blueprint for giants: modeling the physiology of large dinosaurs”. In James O. Farlow; M. K. Brett-Surman. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 491–504. ISBN 978-0-253-21313-6
- ^ Chinsamy, Anusuya; Willem J. Hillenius (2004). “Physiology of nonavian dinosaurs”. In David B. Weishampel; Peter Dodson; Halszka Osmólska. The dinosauria. Berkeley: University of California Press. pp. 643–659. ISBN 978-0-520-24209-8
- ^ Seymour, Roger S. (2013-07-05). “Maximal Aerobic and Anaerobic Power Generation in Large Crocodiles versus Mammals: Implications for Dinosaur Gigantothermy”. PLOS ONE 8 (7): e69361. Bibcode: 2013PLoSO...869361S. doi:10.1371/journal.pone.0069361. ISSN 1932-6203. PMC 3702618. PMID 23861968.
- ^ a b Farlow, James O.; Pianka, Eric R. (2002). “Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs”. Historical Biology 16 (1): 21–40. doi:10.1080/0891296031000154687. オリジナルの2021-11-10時点におけるアーカイブ。 2019年12月2日閲覧。.
- ^ Varricchio, David J. (2001). “Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts”. Journal of Paleontology 75 (2): 401–406. doi:10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2. ISSN 0022-3360.
- ^ Guinness World Records Ltd. (2003). 2003 Guinness World Records. pg 90.
- ^ Strganac, C., Jacobs L., Polcyn M., Mateus O., Myers T., Araújo R., Fergunson K. M., Gonçalves A. O., Morais M. L., Schulp A. S., da Tavares T. S., & Salminen J. (2014). Geological Setting and Paleoecology of the Upper Cretaceous Bench 19 Marine Vertebrate Bonebed at Bentiaba, Angola. Netherlands Journal of Geosciences. 1-16.
- ^ a b Eberth, David A.; McCrea, Richard T. (2001). “Were large theropods gregarious?”. Journal of Vertebrate Paleontology 21 (Supplement to Number 3): 46A. doi:10.1080/02724634.2001.10010852.
- ^ Roach, Brian T.; Brinkman, Daniel L. (2007). “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ a b Currie, Philip J. (1998). “Possible evidence of gregarious behavior in tyrannosaurids”. Gaia 15: 271–277. オリジナルの2009-03-26時点におけるアーカイブ。 2008年9月13日閲覧。.
- ^ Roach, Brian T.; Brinkman, Daniel T. (2007). “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ McCrea, Richard T.; Buckley, Lisa G.; Farlow, James O.; Lockley, Martin G.; Currie, Philip J.; Matthews, Neffra A.; Pemberton, S. George (2014). “A 'Terror of Tyrannosaurs': The First Trackways of Tyrannosaurids and Evidence of Gregariousness and Pathology in Tyrannosauridae”. PLOS ONE 9 (7): e103613. Bibcode: 2014PLoSO...9j3613M. doi:10.1371/journal.pone.0103613. PMC 4108409. PMID 25054328.
- ^ “Tyrannosaurus Tracks Show Dinosaur's Walk” (2014年7月23日). 2014年9月30日時点のオリジナルよりアーカイブ。2015年1月16日閲覧。
- ^ “Tracks Hint at the Social Life of Tyrant Dinosaurs” (2014年7月23日). 2015年1月8日時点のオリジナルよりアーカイブ。2015年1月16日閲覧。
- ^ Titus, Alan L.; Knoll, Katja; Sertich, Joseph J. W.; Yamamura, Daigo; Suarez, Celina A.; Glasspool, Ian J.; Ginouves, Jonathan E.; Lukacic, Abigail K. et al. (April 19, 2021). “Geology and taphonomy of a unique tyrannosaurid bonebed from the upper Campanian Kaiparowits Formation of southern Utah: implications for tyrannosaurid gregariousness”. PeerJ 9: e11013. doi:10.7717/peerj.11013. PMC 8061582. PMID 33976955.
- ^ Eilperin, Juliet (April 19, 2021). “Tyrannosaurs likely hunted in packs rather than heading out solo, scientists find”. The Washington Post. オリジナルのApril 20, 2021時点におけるアーカイブ。 September 27, 2021閲覧。
- ^ “Tyrannosaurus Rex 'hunted in packs'”. 2021年9月27日時点のオリジナルよりアーカイブ。2021年9月27日閲覧。
- ^ a b c Jacobsen, A.R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ^ a b Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ^ Lambe, L. B. (1917). “The Cretaceous theropodous dinosaur Gorgosaurus ”. Memoirs of the Geological Survey of Canada 100: 1–84. doi:10.4095/101672. オリジナルの2021-09-09時点におけるアーカイブ。 2021年9月23日閲覧。.
- ^ Kowalewski, M.: “The Fossil Record of Predation”. pp. 251–266 (2002年). 2008年10月31日時点のオリジナルよりアーカイブ。2022年10月11日閲覧。
- ^ Dorey, M. (1997). Tyrannosaurus. Dinosaur Cards. Orbis Publishing Ltd. D36045907.
- ^ a b Horner, J.R. (1994). “Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger)”. The Paleontological Society Special Publication 7: 157–164. doi:10.1017/S2475262200009497.
- ^ Amos, J. (31 July 2003). “T. rex goes on trial”. BBC. オリジナルの13 March 2012時点におけるアーカイブ。 12 October 2008閲覧。
- ^ a b Walters, M., Paker, J. (1995). Dictionary of Prehistoric Life. Claremont Books. ISBN 1-85471-648-4.
- ^ Erickson G. M.; Olson K. H. (1996). “Bite marks attributable to Tyrannosaurus rex: preliminary description and implications”. Journal of Vertebrate Paleontology 16 (1): 175–178. doi:10.1080/02724634.1996.10011297. オリジナルの2021-10-19時点におけるアーカイブ。 2020年9月12日閲覧。.
- ^ Carpenter, K. (1998). “Evidence of predatory behavior by theropod dinosaurs”. Gaia 15: 135–144. オリジナルの17 November 2007時点におけるアーカイブ。 5 December 2007閲覧。.
- ^ Fowler, Denver W.; Robert M. Sullivan (2006). “A ceratopsid pelvis with toothmarks from the Upper Cretaceous Kirtland Formation, New Mexico: evidence of late Campanian tyrannosaurid feeding behavior”. New Mexico Museum of Natural History and Science Bulletin 35: 127–130.
- ^ Longrich, N. R.; Horner, J. R.; Erickson, G. M.; Currie, P. J. (2010). “Cannibalism in Tyrannosaurus rex”. PLOS ONE 5 (10): e13419. Bibcode: 2010PLoSO...513419L. doi:10.1371/journal.pone.0013419. PMC 2955550. PMID 20976177.
- ^ “NEW EVIDENCE FOR CANNIBALISM IN TYRANNOSAURID DINOSAURS FROM THE UPPER CRETACEOUS (CAMPANIAN/MAASTRICHTIAN) SAN JUAN BASIN OF NEW MEXICO”. ResearchGate. 2021年11月10日時点のオリジナルよりアーカイブ。2021年1月6日閲覧。
- ^ Dodson, Peter; Britt, Brooks; Carpenter, Kenneth; Forster, Catherine A.; Gillette, David D.; Norell, Mark A.; Olshevsky, George; Parrish, J. Michael et al. (1993). “Abelisaurus”. Age of Dinosaurs. Publications International, LTD. p. 105. ISBN 978-0-7853-0443-2
- ^ Fiorillo, Anthony R.; Gangloff, Roland A. (2000). “Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on arctic dinosaur paleoecology”. Journal of Vertebrate Paleontology 20 (4): 675–682. doi:10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2. ISSN 0272-4634.
- ^ “国内初のティラノサウルス科大型種の歯の化石について”. 福井県立恐竜博物館 (2015年7月14日). 2022年10月6日閲覧。
- ^ Thomas R. Holtz Jr. (1994). “The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics”. Journal of Paleontology 68 (5): 1100–1117. doi:10.1017/S0022336000026706. JSTOR 1306180.
- ^ February 2016, Laura Geggel 29 (29 February 2016). “T. Rex Was Likely an Invasive Species”. livescience.com. 14 July 2019時点のオリジナルよりアーカイブ。5 January 2021閲覧。