
アノマロカリス類
| アノマロカリス類 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| ||||||||||||||||||
| 地質時代 | ||||||||||||||||||
| カンブリア紀(Cambrian Stage 3)[1] - デボン紀前期[2] | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Radiodonta Collins, 1996 | ||||||||||||||||||
| 英名 | ||||||||||||||||||
| Anomalocaridid Anomalocarid Radiodontid Radiodontan Radiodont | ||||||||||||||||||
| 属 | ||||||||||||||||||
|
アノマロカリス類(Radiodont、Radiotontan、Anomalocaridid)は、基盤的な節足動物と考えられる古生物の1グループ、放射歯目(ラディオドンタ目、Radiodonta)に該当する総称である。ラディオドンタ類とも呼ばれ[7]、アノマロカリスだけでなく、アンプレクトベルア、ペユトイア、およびフルディアなどをも含んだ多様な分類群である。
関節に分かれる前部付属肢・頭部を覆う甲皮・眼柄に備わる複眼・胴部に配置される鰓と対をなしに並んだ鰭(ヒレ)という独特な特徴の組み合わせをもつ海棲動物である。カンブリア紀の大型動物として知られる種類を中心とし、その習性は活発な肉食性から穏やかな濾過摂食性まで多岐にわたる[8][9]。
世界中の化石産地から見つかっており、カンブリア紀における代表的な古生物であるが、オルドビス紀とデボン紀に生息した種類ものちに発見される[2][5]。その形態の復元と分類学上の位置付けは多くの議論が繰り広げられ、節足動物の起源と初期系統発生を示唆するのに重要視される分類群の1つである。
学名と総称
時期と文献によって、アノマロカリス類に採用した分類階級や該当する学名の定義はやや異なる。それに応じて、英語には「anomalocaridid[5][10][11][4]」・「anomalocarid[11]」・「radiodontid[12]」・「radiodontan[6][13][14]」・「radiodont[8][15]」など多くの総称が用いられていた。
- アノマロカリス科 Anomalocarididae Raymond, 1935
- アノマロカリス亜目 Anomalocarida Vinther et al., 2014[11]
- 総称「anomalocarid」の由来。Vinther et al., 2014 によって創設され、4つの科を含んだ系統群。後述も参照。
- 放射歯目[16](ラディオドンタ目[7])Radiodonta Collins, 1996[12]
形態


体長が30-50cm程度の大型種を中心とするグループで、1mを超える超大型種や数cmしか及ばない小型種も僅かに発見される[8]。既知最大の属は2mほどの巨体をもつエーギロカシス属で、最小の属は体長8cm程度のライララパクス属であり[8]、後者の1種 Lyrarapax unguispinus の幼生に至っては体長は1.8cmしか及ばない。
上下に扁平または円柱状に近い体型をもつ[17]。体節制があり、前後で大まかに先節のみからなる頭部と数節-十数節からなる胴部として区別できる。ほとんどの表皮(クチクラ)は柔軟で、真の節足動物のように硬質化していないと考えられる[13]。また、ククメリクルスを除いて、腹側には脚およびその接続部らしい構造が見当たらない[3]。
アノマロカリス類は、基本として以下の特徴の組み合わせで他の動物から区別できる[9]。


- 頭部:
- 胴部:
頭部の甲皮

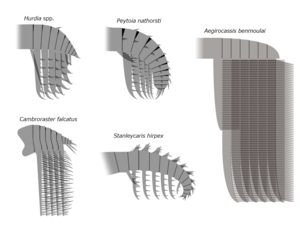
アノマロカリス類の頭部には甲皮のような硬組織があり、「head shield」・「cephalic carapace」・「head sclerite」・「head carapace complex」など様々な通称で呼ばれる。これは基本として3つのパーツからなり、中心の1枚(H-element、dorsal carapace、dorsal plate、anterior sclerite)は頭部の背側、残りの2枚(P-elements、lateral sclerites)は頭部の左右に備わり、前端の突出部(P-element neck、beak)を通じて連結している場合がある[6][18][19][9]。かつて、このような構造体はフルディアのみに備わると考えられてきたが、後に再検証や新たな化石証拠で他の多くのアノマロカリス類にも見られるようになり、本群全般に共有の形質であると分かった。
遊泳性の獲物を摂る活発な捕食者であったと思われるアノマロカリスとアンプレクトベルア科の頭部の甲皮は小さく、捕獲用の前部付属肢の可動範囲の邪魔にならない同時に防衛の役割をも果たしていたと思われる[18]。一方で、底生性で堆積物から食物を摂ると考えられるフルディアとカンブロラスター、および水中のプランクトンを摂る濾過摂食者とされるエーギロカシスなど多くのフルディア科の種類は、頭部の甲皮が著しく発達しており、巨大な構造体になっている[5][18]。
前部付属肢


アノマロカリス類の代表的な特徴である "触手" や "触腕" は前部付属肢[20](Frontal appendages)といい、他にも「feeding appendages」・「grasping appendages」などと呼ばれ[10]、Megacheiraに近縁とされた時期では「great appendages」(大付属肢)扱いともされてきたが、両者は類縁でない別系統で、前部付属肢と大付属肢は非相同の付属肢であると判明して以降、この総称をアノマロカリス類の前部付属肢に用いられなくなるようになった。
前部付属肢はアノマロカリス類の唯一の関節肢であり、口の前方に左右2本を持ち、多数の肢節(podomere)に分かれ、上下(カリョシントリプスの場合は左右)に運動でき、造形はエビの腹部に似ている。多くの場合、前部付属肢は前後で基部1-3節からなる柄のような部分(shaft)と残りの数節-十数節からなる部分(distal articulated region)として区別でき、腹側は多くの節ごとに1本もしくは1対の内突起(endite)がある[21][22]。硬質で、様々な器官のうちこの付属肢は往々にして保存状態が最も良好な部分であるため、アノマロカリス類の中で、前部付属肢のみによって知られる種類は少なくない[23]。
-
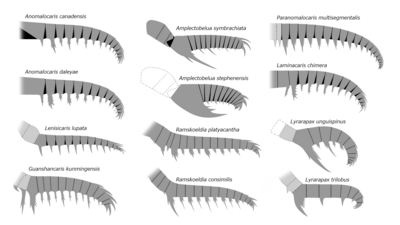 アノマロカリス科、アンプレクトベルア科、およびそれらに類する種の前部付属肢。
アノマロカリス科、アンプレクトベルア科、およびそれらに類する種の前部付属肢。 -
 タミシオカリス科の前部付属肢。
タミシオカリス科の前部付属肢。 -
 フルディア科の前部付属肢。
フルディア科の前部付属肢。 -
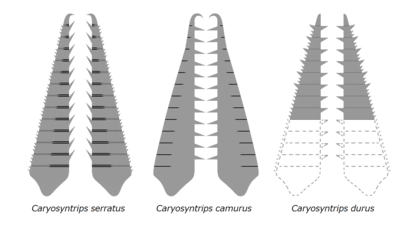 カリョシントリプスの前部付属肢。
カリョシントリプスの前部付属肢。




前部付属肢の形態、特に内突起は種によって構造が異なり、同定形質、およびその動物の食性と生態を示唆する部分として重要視される[24][25]。例えばアノマロカリスのは運動性の高い触手状で、アンプレクトベルアとライララパクスのは鋏のように嚙合わせた大きな内突起があり、ペユトイアとフルディアのは短い熊手状で5本のブレード状の内突起を持ち、タミシオカリスとエーギロカシスのは懸濁物食や濾過摂食に適した繊毛状の構造を内突起に並んでいる[5]。腹側の節間膜の面積から、その可動範囲を推測することもできる[11][23]。
ライララパクスの化石に保存された神経系の痕跡により、前部付属肢の神経は前大脳の前方に対応(前大脳性)であると判明した。従ってこの付属肢は先節由来で、有爪動物の触角と真節足動物の上唇に相同であり、他の節足動物の鋏角、第1触角、および大付属肢(いずれも中大脳性で第1体節由来)には相同でないと考えられる[4]。
口(歯)


「Oral cone」と呼ばれる円盤状の口器。頭部の下面、前部付属肢の付け根の後方にあり、往々にしては放射状に並んだ歯によって構成され、パイナップルの輪切りのような造形をもつ。そのうち最も発達した歯は4枚で典型的な「ペユトイア口」の十字放射(フルディア科、ライララパクス)、または3枚で三放射(アノマロカリス)に並んでいるものが知られる[3]。フルディアとCambrorasterの場合、開口部の奥にはノコギリ状の多重構造が加えている[17][9]。アンプレクトベルアとRamskoeldiaの場合、歯の構造は不明確であるが、放射状でないと考えられる[6][15]。
「放射状の歯」を意味するアノマロカリス類の学名「Radiodonta」に表れるように[25]、かつて、このような口はアノマロカリス類と環神経動物(汎節足動物以外の脱皮動物)に特有の形質と考えられたが、アノマロカリス類の近縁であるパンブデルリオンは放射状に並んだ歯を持ち[26]、他の葉足動物からにもらしき口器が次々と発見される。そのため、この放射状の歯は後にアノマロカリス類と環神経動物に限る派生形質ではなく、むしろ脱皮動物全般の祖先形質とされるようになった(葉足動物#口器も参照)[27]。
Setal blades

アノマロカリス類のそれぞれの体節の背面には、「setal blades[5]」(または「lamellae[9]」)と呼ばれる櫛状の構造体がある。これは無数の細長い葉状附属体「lanceolate blades」から構成されており、管状の内部構造がそれぞれのlanceolate bladesの前縁部付近を貫通しながら繋がっていたと思われる[5]。この部分は、中心から左右に区切れて対になるものと、一面の構造体になって左右が完全に会合するものがある。背側のヒレが存在する場合、この構造の両端はその基部に接続していたと考えられる[5]。機能に関しては呼吸用の鰓であった可能性が多くの文献に挙げられており[3]、一部の種に見られ、その表面にある表面積を増やした皺構造もこの仮説に裏付ける[5]。この構造体は本群の近縁とされるオパビニアにも見られるが、胴部の背側の代わりにヒレの表面に張り付くという相違点がある[5]。パンブデルリオンとケリグマケラのヒレの表面にある繊毛状の構造は、この部位に相同であると考えられる[5]。
多くのアノマロカリス類の部位と同様、これも遺骸ないし脱皮殻から脱落しやすい部分であると考えられる[28]。フルディアのはよく発達しているため化石例が少なからず見つかっており[28]、代わりにアノマロカリス属やアンプレクトベルア科においてこの部位が保存された化石は非常に稀である[6]。多くのアノマロカリス類の復元図において、この部分は往々にして描かれていない。
鰭(ヒレ)


「Flaps」ないし「lobes」といい、付属肢に由来で遊泳用の器官であると考えられる。腹側から左右に向かって張り出している。多くの種類には、ヒレの基部は次のヒレに覆われる部分があり、後方から1対ずつ重なり合う構造をもつ。また、ヒレは往々にして筋のような構造が見られ、「strengthening rays」・「tranverse rods」・「veins」などと呼ばれ、ヒレを支える骨幹として機能をする部分と思われる[3]。
通常は1体節に腹側からの1対のみ発見されているが、エーギロカシスとペユトイアなどフルディア科のものは、背側から退化的なヒレ(dorsal flaps)の存在が確認されており、その付け根はSetal bladesに接続していた(この場合、腹側のヒレは「ventral flaps」と呼ぶ)。この発見により、アノマロカリス類の基本体制および他の汎節足動物の付属肢対応関係が書き替えられた(エーギロカシス#発見の意義を参照)[5]。
多くの種類は体に並ぶ大きなヒレの他に、頭部の付近、いわゆる"首"に当たる部分にも3-4対の退化的なヒレ(head flaps、anterior flaps)が並んでいる[3][17]。アンプレクトベルアと Ramskoeldia の場合、この退化的なヒレの付け根には「gnathobase-like structure」(GLS)という顎基らしい構造をもつ[6][15]。
尾部

多くのアノマロカリス類の胴部の後端には、「tail fan」および「tail fin」と呼ばれ、対をなしに特殊化したヒレ状の付属体がある。アノマロカリス・カナデンシスは3対、アノマロカリス・サロンとCambrorasterは2対、フルディアとシンダーハンネスは1対、ペユトイアとエーギロカシスは尾びれを持たず、尾端は単純の鈍い突起である[5]。少なくともアノマロカリス・カナデンシスの場合、第3対の尾びれの間にはもう1本の小さなヒレらしい尾状突起がある[3]。アンプレクトベルアとライララパクスでは、1対の尾毛がある[9]。シンダーハンネスは、尾びれの次には1本の剣状の尾がある[2]。アノマロカリス類の近縁とされるオパビニアとケリグマケラも、それぞれ特殊化した3対の尾びれと1本の剣状の尾がある。
内部構造

内部の器官は、消化管、筋肉組織と神経系が発見されている。消化管は他のステムグループ節足動物(シベリオン科とGilled lobopodians)や基盤的な真節足動物(フキシャンフィア類、イソキシスなど)のように、中腸は体節の境目に応じて6対の分岐(消化腺、中腸腺)が並んでいる[3][29][30]。それより外縁の領域ではヒレの数に応じた筋組織が詰まっている[3][4]。神経系はライララパクスの化石から最初に発見され、有爪動物に似通う単純なはしご形神経系である。脳の神経節は前大脳1つだけで、前部付属肢と複眼の神経はそれぞれ前大脳の前部と両側に繋がっている[4]。
生態
生理学

アノマロカリス類は自由遊泳の生活をする動物(ネクトン)であったと思われる。発達の筋肉を有する体のヒレを波打たせることにより水中で推進できたと推測され、この動作は、現生のエイや、コウイカの仲間に似通うと考えられる[31]。一部の種類に見られる背側のヒレや尾端の尾びれの役割に関しては、遊泳の動作を安定させるために使っており[2]、飛行機の尾翼のように横安定性を維持し、鳥類の尾羽のようにターンを行う効率を上げる機能をもつ、などの説がある[32]。また、脱落した硬組織の化石が群れに発見される例が少なからぬ挙げられることにより、アノマロカリス類は一部の節足動物のように、集団で脱皮を行ったことが示唆される[9]。
食性など




かつては全般的に獰猛な捕食者であったと考えられたが、アノマロカリス類の生態は必ずしもそうとは限らず、多様だったと考えられる。これは前部付属肢の形で明瞭に示唆されており、捕食性に適したアノマロカリス属の触手状とアンプレクトベルア属の鋏状から[24]、堆積物食に向いているペユトイアとフルディアの熊手状のものや[24][33]、エーギロカシスとタミシオカリスのような懸濁物食や濾過摂食に適した櫛状まで多岐していた[5][11][8]。アノマロカリス類の食性は歯によって示唆されることもあり、例えばアノマロカリスの不規則な三放射状の歯は活発な小型獲物を捕食するのに適しており[33]、多重構造と大きな開口部をもつフルディアとカンブロラスターの歯は堆積物から広い範囲の食物を摂食できたと考えられる[9]。


体型・ヒレ・眼・甲皮など種によって各部位の相違点も、本群の生態と当時の食物連鎖における役割の多様性を示唆する[8]。代表的な例として、アノマロカリス属とペユトイア属(ラガニア属)を比較すると、アノマロカリスは流線型の体・発達した尾部のヒレ・高い可動域をもつ長い前部付属肢を有し、複眼も頭部の前端付近に備わり、良好な視覚を有する活発な遊泳性の捕食者であったことを示唆する[34]。一方で、ペユトイアではくびれの無い楕円形の体型と短い熊手状の前部付属肢を有し、尾部のヒレを欠いており、眼は頭部の後方にあった。これらの特徴は能動的な捕食について不利であり、底生性で堆積物から餌を摂ったと考えられる[33]。他にも、多くのアノマロカリス類とは対照的に小型であるライララパクスや、ドーム型の甲皮により更に底生性の生態に適したカンブロラスターなどが挙げられる[9]。
発見史
アノマロカリス類のほとんどの表皮は柔軟で、硬質化した部位も局部に限られるため、遺骸と脱皮殻の各部分はばらばらになりやすい[28]。そのため、良好な保存状態をもつ全身化石が残ることは非常にまれであり、散在した部位は、しばしばアノマロカリス類でない独立した別の生物やその一部と誤解され、もしくは逆に複数の種のアノマロカリス類の特徴を誤って1つの種に由来とされることがあった。

-

-
 前部付属肢と共に保存される同種の全身化石。
前部付属肢と共に保存される同種の全身化石。 -
 同種の全身復元図。
同種の全身復元図。



著名な例として、アノマロカリス・カナデンシスとペユトイア・ナトルスティが挙げられる。この2種のアノマロカリス類の化石は最初では局部か散在した状態で発見され、前者は前部付属肢のみで、「アノマロカリス」(アノマロカリス・カナデンシス Anomalocaris canadensis)というコノハエビ類の腹部として記載されており[12]、この「甲殻類の腹部」と誤解されていた化石が、つねに頭部を欠いているのが謎とされていた。後者の歯の部分はクラゲの化石と考えられ、「ペユトイア」(ペユトイア・ナトルスティ Peytoia nathorsti)と記載される同時に、その胴体は海綿かナマコの一種と見なされ、「ラガニア」(ラガニア・カンブリア Laggania cambria)と名付けられた[35]。これらの化石は、最初はそれぞれが関係しているとは考えられていなかった。これらの部位を1つの動物体に出揃った全身化石が発見され、全身復元が再構成された1980年代以降、何種類かの属や種が発見記載された。それらは、前部付属肢の細部、尾部のヒレの有無、歯の配置、その他の特徴に違いが見られる[36]。また、アノマロカリス類として再構成されるうちに、ある種のものとされた部位は、実は別の種類のものであり、散在した化石によって混同されることがその後に判明したものもあり、例えばかつてアノマロカリスのものと考えられた歯の化石はペユトイアに由来のもので[37]、フルディアのものと考えられてきた一部の甲皮の化石は Pahvantia に由来のものであると後に分かった[8]。
「奇妙なエビ」を表す「Anomalocaris」(アノマロカリス)という名称は、もともとちぎれた前部付属肢に対して付けられた名称だったが、その後この動物体全体の名称として使用された。歯と胴部がそれぞれ別の生物とされたペユトイアの場合は学名の先取権の原則によるもので、歯の部分である「ペユトイア」をタイプとし、胴部を示す「ラガニア」はそのシノニムとされる[37]。
生息時代
アノマロカリス類は、主にカンブリア紀の地層から産出する化石によって知られ、中国、アメリカ、カナダ、ポーランド、オーストラリアなどで発見されている。なお、これらはいずれも既知の最古の三葉虫化石と真節足動物の生痕化石に先んじるものであり、アノマロカリス類と真節足動物の間には中間的化石が欠如していることを示唆する[1]。
アノマロカリス類の体は化石に残りにくく、中間の期間の化石は残されていない。長い間、カンブリア紀の中期と後期の間(ミドルカンブリアンの終焉期)を境に長らく化石が発見されなかったため、本群の生息期間はカンブリア紀に限定され、そこで絶滅したと考えられていた。その時期には地球環境に大きな変化は無く、温暖な気候が続いていたと考えられているため、なぜ突如として絶滅したのかは謎とされていた。
しかし2009年、デボン紀の地層からアノマロカリス類と考えられるシンダーハンネスの化石が発見されるに至り[2]、アノマロカリス類はカンブリア紀を超えて生存していたことが分かり、生息期間の記録が数千万年も延長された。2011年5月、モロッコで新たに、いくつかのアノマロカリス類の軟組織を残したオルドビス紀の化石が見つかり[38]、注目を集めている。更に2015年、前述の一部の化石は、フルディアによく似た全長2メートルの大型アノマロカリス類のものであると判明し、エーギロカシス(Aegirocassis)として記載された[5]。
分類
系統関係
| |||||||||||||||||||||||||||||||||||||||||||||
| 脱皮動物におけるアノマロカリス類の系統位置[5][11][28]。 †:絶滅群 *:Dinocarididaに分類される群 |






アノマロカリス類は、オパビニア、パンブデルリオンやケリグマケラなどと共に、節足動物が葉足動物から一歩ずつ分化する過程に当たる、その系統発生の様子を推定する重要な動物群として注目される。これらの動物群はいずれも一連のヒレ状付属肢と特殊化した前部付属肢という共有形質が見られ、Dinocaridida綱[12]というステムグループ節足動物(節足動物の初期脇道系統)の1群としてまとめられる。一見では節足動物に類似しないが、いずれも基盤的な真節足動物らしい消化腺をもち[29]、特にアノマロカリス類は、関節肢(前部付属肢)と複眼など節足動物との重要な共有形質が出揃っている。アノマロカリス類の背腹2対のヒレや、パンブデルリオンなどの同時に葉足とヒレを持った体制も、節足動物の二叉型付属肢の起源を示唆する形質と見なされる[5]。アノマロカリス類の頭部における背側の甲皮も、フキシャンフィア類やオダライアなどの基盤的な真節足動物の頭部における小さな外骨格と同様に複眼と前大脳性(先節由来)であるため、相同性が挙げられる[19]。
これらの動物は、節足動物のステムグループに属する基盤的な節足動物と考えられ、そのうちアノマロカリス類、オパビニアと真節足動物はお互いの姉妹群とする知見が広く認められる[11][39][40]。
他にもMegacheira類に類縁と考え、お互いの大付属肢/前部付属肢を相同器官と見なし、共に鋏角類のステムグループに属する[41]、もしくは放射状の歯に基づいて、汎節足動物以外の脱皮動物(環神経動物 Cycloneuralia)に類縁する[10]などの異説はかつてあったが、いずれものちに多くの研究に否定的とされる[5][4]。前者は、神経解剖学的証拠に基づいてお互いの大付属肢/前部付属肢は別の体節に由来の相似器官であると判明した[4]。後者の基準となった放射状の歯は、あくまでも脱皮動物の共有原始形質にすぎず[27]、はしご形神経系の発見も、アノマロカリス類と環神経動物の類縁関係を否定し、汎節足動物であることを示唆する強力な証拠となる[4]。また、シンダーハンネスのいくつかの真節足動物的とされる形質に基づいて、アノマロカリス類を真節足動物に至る側系統群と見なす見解はかつてあった[2]が、再検討によりその形質は曖昧で真節足動物らしきぬとされ、この説も否定的とされるようになった[13](該当記事を参照)。
近縁


近縁の古生物として、上述のオパビニア(Opabinia)、パンブデルリオン (Pambdelurion)とケリグマケラ (Kerygmachela)がある。いずれもアノマロカリス類と同様にDinocaridida綱に属とされ、一連のヒレと特殊化した前部付属肢を持ち、「Gilled lobopodians」(鰓のある葉足動物)とも呼ばれた動物群である。アノマロカリス類に似通う特徴は他にもいくつか見られ、例えばオパビニアの特殊化した尾ヒレにあること・「setal blades」をもつこと・眼が眼柄にあることが本群と共通し、パンブデルリオンは特殊化した放射状の歯をもち[26]、ケリグマケラは複眼様の多数のレンズからなる眼がある[42]。
これらの動物のアノマロカリス類との主な相違点は、胴部の前端数節が"首"になれず[9]、前部付属肢は左右開閉で関節肢化せず、アノマロカリス類とは逆の畳み方を持ったヒレ、鰓らしき櫛状構造「setal blades」がヒレの表面に張り付き、腹側には脚(葉足)が有する、などの特徴が挙げられる[5]。これらの動物は、葉足動物の共有原始形質を色濃く備えており、アノマロカリス類より基盤的な系統から派生した群であると見なされる[5]。
疑問視される種
-
カリョシントリプスの前部付属肢は左右開閉の構造をもつ。
-
 嚢頭類の甲皮の片割れとして復元されたZhenghecaris。
嚢頭類の甲皮の片割れとして復元されたZhenghecaris。

カリョシントリプス(Caryosyntrips)とククメリクルス(Cucumericrus)は、通常ではアノマロカリス類と見なされる[5][24][23][43]。しかしカリョシントリプスの前部付属肢は、左右開閉の構造をもつなどアノマロカリス類との相違点が見られており[44]、それ以外の部位も未だに発見されていない。ククメリクルスの前部付属肢は未だに発見できず、ヒレの下面には脚を具えており、しかも葉足と関節肢の両方に似通う中間的形態がみられる[45]。全身化石が出揃っていないこれらの古生物はアノマロカリス類として記載されるものの、アノマロカリス類としての形質がしばしば疑問視される。Zhenghecaris という左右相称のドーム状の外骨格のみによって知られる化石は、原記述では嚢頭類(Thylacocephala)の甲皮と考えられてきたが、カンブロラスターらしきフルディア科のアノマロカリス類の甲皮である可能性がのちに取り上げられる[9]。
下位分類
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Aubril et al. (2018) に基づいたアノマロカリス類の系統関係[8]。正式の記載がなされていない化石はここで省略される。 |

前述の疑問種をも含め、アノマロカリス類とされるものは以下の属が知られている。
- アノマロカリス Anomalocaris [36]
- ペユトイア Peytoia(旧ラガニア Laggania)[37]
- フルディア Hurdia [28]
- アンプレクトベルア Amplectobelua [10]
- ? ククメリクルス Cucumericrus[10]
- シンダーハンネス Schinderhannes [2][11]
- カリョシントリプス Caryosyntrips [24]
- スタンレイカリス Stanleycaris [46]
- タミシオカリス Tamisiocaris [11]
- パラノマロカリス Paranomalocaris [47]
- ライララパクス Lyrarapax [4]
- エーギロカシス Aegirocassis [5]
- Ramskoeldia [15]
- ラミナカリス[7] Laminacaris [25]
- Pahvantia [8]
- Ursulinacaris [22]
- カンブロラスター[7] Cambroraster [9]
- ? Zhenghecaris [9]
かつて、全てのアノマロカリス類はアノマロカリス科(Anomalocarididae)に分類された。新たな分類体系が Vinther et al., 2014 に創設されて以降、アノマロカリス類は放射歯目(Radiodonta)に当たる群とされるようになり、そのうち多くの種類がアンプレクトベルア科(Amplectobeluidae)・タミシオカリス科(Tamisiocarididae、旧Cetiocaridae)・フルディア科(Hurdiidae)という新たに創設される3つの科に再分類された[48]。カリョシントリプスをこの類に含まれる場合、本属はどの科にも属さず、残り全てのアノマロカリス類(アノマロカリス亜目 Anomalocarida)の姉妹群として基盤的なアノマロカリス類とされる[5][11][8]。アノマロカリス類の中で、フルディア科は最も多様化しており、2019現在では9つの属が本科に含まれる[9]。
アノマロカリス類の内部系統関係について、フルディア科の単系統性は広く認められるが、アノマロカリス科とアンプレクトベルア科の内部構成は不確実である[25]。特にアノマロカリス科のアノマロカリス属は往々にして非単系統群であるとされ、分岐学的にはタイプ種であるアノマロカリス・カナデンシスよりも、別のアノマロカリス類(主にアンプレクトベルア科)に近い系統に位置付けされる種類がほとんどである[8][4][48]。ライララパクスは従来ではアンプレクトベルア科に分類されるが、歯と首の構造の相違点によって本科としての形質がしばしば疑問視される[15][25]。
- 放射歯目 Radiodonta
- ? ククメリクルス Cucumericrus
- カリョシントリプス Caryosyntrips
- アノマロカリス亜目 Anomalocarida
- パラノマロカリス Paranomalocaris(一部の文献ではアノマロカリス科に分類される[5][49])
- ラミナカリス Laminacaris (一部の文献ではアンプレクトベルア科に分類される[8])
- アノマロカリス科 Anomalocarididae
- アノマロカリス Anomalocaris(一部の種は別系統とされる場合がある[8])
- アンプレクトベルア科 Amplectobeluidae
- アンプレクトベルア Amplectobelua
- ライララパクス Lyrarapax(異説あり[15][25])
- Ramskoeldia(異説あり[9])
- "アノマロカリス"・クンミンゲンシス "Anomalocaris" kunmingensis
- タミシオカリス科 Tamisiocarididae(旧Cetiocaridae)
- タミシオカリス Tamisiocaris
- "アノマロカリス"・ブリッグシ "Anomalocaris" briggsi
- フルディア科 Hurdiidae
本群とは見なされなかった古生物
限られた部位によって知られ、それをアノマロカリス類に由来と誤解された別生物は、以下の例が挙げられる。

- 付属肢と腹板のみによって知られる。1995年に「脚のあるアノマロカリス類」として復元されたが、2010年代以降からMegacheira類の真節足動物と見なされるようになった[50][51][52][3][5][53]。(詳細はパラペユトイアを参照)
- 一連の歯からなる口器のみによって知られる。1994年に巨大なアノマロカリス類のoral coneと考えられ[55]、2008年に鰓曳動物の口器とされると同時に新属新種として記載された[54]が、2016年にパンブデルリオンらしい動物の口器と見なされるようになった[26]。
脚注
- ^ a b Daley, Allison; Antcliffe, Jonathan; Drage, Harriet; Pates, Stephen (2018-05-21). “Early fossil record of Euarthropoda and the Cambrian Explosion”. Proceedings of the National Academy of Sciences 115: 201719962. doi:10.1073/pnas.1719962115.
- ^ a b c d e f g Kühl, G.; Briggs, D. E. G.; Rust, J. (Feb 2009). “A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany”. Science 323 (5915): 771–3. doi:10.1126/science.1166586. ISSN 0036-8075. PMID 19197061.
- ^ a b c d e f g h i j k C., Daley, Allison; D., Edgecombe, Gregory. “Morphology of Anomalocaris canadensis from the Burgess Shale” (英語). Journal of Paleontology 88 (01). ISSN 0022-3360.
- ^ a b c d e f g h i j k l Peiyun Cong; Xiaoya Ma; Xianguang Hou; Gregory D. Edgecombe; Nicholas J. Strausfeld (2014). “Brain structure resolves the segmental affinity of anomalocaridid appendages”. Nature 513 (7519): 538–42. doi:10.1038/nature13486. PMID 25043032.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2015). “Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps”. Nature 522 (7554): 77–80. doi:10.1038/nature14256. ISSN 0028-0836. PMID 25762145.
- ^ a b c d e f g h Cong, Peiyun; Daley, Allison C.; Edgecombe, Gregory D.; Hou, Xianguang (2017-08-30). “The functional head of the Cambrian radiodontan (stem-group Euarthropoda) Amplectobelua symbrachiata” (英語). BMC Evolutionary Biology 17 (1). doi:10.1186/s12862-017-1049-1. ISSN 1471-2148. PMC PMC5577670. PMID 28854872.
- ^ a b c d 土屋, 健 (サイエンスライター) (2020-2-12). アノマロカリス解体新書. 田中 源吾 (監修), かわさき しゅんいち (イラスト). 東京: ブックマン社. ISBN 978-4-89308-928-1. OCLC 1141813539
- ^ a b c d e f g h i j k l m Lerosey-Aubril, Rudy; Pates, Stephen (2018-09-14). “New suspension-feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton” (英語). Nature Communications 9 (1). doi:10.1038/s41467-018-06229-7. ISSN 2041-1723.
- ^ a b c d e f g h i j k l m n o p q Moysiuk J.; Caron J.-B. (2019-08-14). “A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources”. Proceedings of the Royal Society B: Biological Sciences 286 (1908): 20191079. doi:10.1098/rspb.2019.1079.
- ^ a b c d e f Xian‐Guang, Hou; Bergström, Jan; Ahlberg, Per (1995-09-01). “Anomalocaris and Other Large Animals in the Lower Cambrian Chengjiang Fauna of Southwest China”. GFF 117: 163–183. doi:10.1080/11035899509546213.
- ^ a b c d e f g h i j Jakob Vinther, Martin Stein, Nicholas R. Longrich & David A. T. Harper (2014). “A suspension-feeding anomalocarid from the Early Cambrian”. Nature 507: 496–499. doi:10.1038/nature13010. PMID 24670770.
- ^ a b c d Collins, D. (1996). “The "Evolution" of Anomalocaris and Its Classification in the Arthropod Class Dinocarida (nov.) and Order Radiodonta (nov.)”. Journal of Paleontology 70 (2): 280–293. doi:10.2307/1306391. JSTOR 1306391.
- ^ a b c d Javier, Ortega-Hernández,. “Making sense of ‘lower’ and ‘upper’ stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848” (英語). Biological Reviews 91 (1). ISSN 1464-7931.
- ^ a b Pates, Stephen; Daley, Allison C. (2017-08). “Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada” (英語). Papers in Palaeontology 3 (3): 461–470. doi:10.1002/spp2.1084. ISSN 2056-2802.
- ^ a b c d e f g Cong, Pei-Yun; Edgecombe, Gregory D.; Daley, Allison C.; Guo, Jin; Pates, Stephen; Hou, Xian-Guang (2018-06-23). “New radiodonts with gnathobase-like structures from the Cambrian Chengjiang biota and implications for the systematics of Radiodonta” (英語). Papers in Palaeontology. doi:10.1002/spp2.1219. ISSN 2056-2802.
- ^ “脳構造から明らかになるアノマロカリス類の付属肢の体節との関連性 | Nature | Nature Research”. www.natureasia.com. 2018年9月17日閲覧。
- ^ a b c d Daley, Allison; Budd, Graham; Caron, Jean-Bernard (2013-10-01). “The morphology and systematics of the anomalocarid Hurdia from the Middle Cambrian of British Columbia and Utah”. Journal of Systematic Palaeontology 11. doi:10.1080/14772019.2012.732723.
- ^ a b c d Zeng, Han; Zhao, Fangchen; Yin, Zongjun; Zhu, Maoyan (2017-01-04). “Morphology of diverse radiodontan head sclerites from the early Cambrian Chengjiang Lagerstätte, south-west China”. Journal of Systematic Palaeontology 16: 1. doi:10.1080/14772019.2016.1263685.
- ^ a b “Homology of Head Sclerites in Burgess Shale Euarthropods” (英語). Current Biology 25 (12): 1625–1631. (2015-06-15). doi:10.1016/j.cub.2015.04.034. ISSN 0960-9822.
- ^ “カンブリア紀の「優しい巨人」 | Nature ダイジェスト | Nature Research”. www.natureasia.com. 2019年1月19日閲覧。
- ^ Pates, Stephen; Daley, Allison C. (2019/07). “The Kinzers Formation (Pennsylvania, USA): the most diverse assemblage of Cambrian Stage 4 radiodonts” (英語). Geological Magazine 156 (7): 1233–1246. doi:10.1017/S0016756818000547. ISSN 0016-7568.
- ^ a b Pates, Stephen; Daley, Allison C.; Butterfield, Nicholas J. (2019-12). “First report of paired ventral endites in a hurdiid radiodont” (英語). Zoological Letters 5 (1). doi:10.1186/s40851-019-0132-4. ISSN 2056-306X. PMC PMC6560863. PMID 31210962.
- ^ a b c Pates, Stephen; Daley, Allison C. (2017-08). “Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada” (英語). Papers in Palaeontology 3 (3): 461–470. doi:10.1002/spp2.1084. ISSN 2056-2802.
- ^ a b c d e DALEY, ALLISON C.; BUDD, GRAHAM E. (2010-07-19). “New anomalocaridid appendages from the Burgess Shale, Canada” (英語). Palaeontology 53 (4): 721–738. doi:10.1111/j.1475-4983.2010.00955.x. ISSN 0031-0239.
- ^ a b c d e f g Guo, Jin; Pates, Stephen; Cong, Peiyun; Daley, Allison; Edgecombe, Gregory; Chen, Taimin; Hou, Xianguang (2018-08-17). “A new radiodont (stem Euarthropoda) frontal appendage with a mosaic of characters from the Cambrian (Series 2 Stage 3) Chengjiang biota”. Papers in Palaeontology. doi:10.1002/spp2.1231.
- ^ a b c Vinther, Jakob; Porras, Luis; Young, Fletcher; Budd, Graham; Edgecombe, Gregory (2016-09-01). “The mouth apparatus of the Cambrian gilled lobopodian Pambdelurion whittingtoni”. Palaeontology. doi:10.1111/pala.12256.
- ^ a b Smith, Martin; Caron, Jean-Bernard (2015-07-02). “Hallucigenia’s head and the pharyngeal armature of early ecdysozoans”. Nature 523: 75–78. doi:10.1038/nature14573.
- ^ a b c d e Daley, Allison C.; Budd, Graham E.; Caron, Jean-Bernard; Edgecombe, Gregory D.; Collins, Desmond (2009-03-20). “The Burgess Shale Anomalocaridid Hurdia and Its Significance for Early Euarthropod Evolution” (英語). Science 323 (5921): 1597–1600. doi:10.1126/science.1169514. ISSN 0036-8075. PMID 19299617.
- ^ a b Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. (2014-05-02). “Sophisticated digestive systems in early arthropods” (英語). Nature Communications 5 (1). doi:10.1038/ncomms4641. ISSN 2041-1723.
- ^ Cong, Peiyun; Daley, Allison (英語). Morphology of the Radiodontan Lyrarapax from the Early Cambrian Chengjiang Biota.
- ^ Usami, Yoshiyuki (2006-01-07). “Theoretical study on the body form and swimming pattern of Anomalocaris based on hydrodynamic simulation”. Journal of Theoretical Biology 238 (1): 11–17. doi:10.1016/j.jtbi.2005.05.008. ISSN 0022-5193. PMID 16002096.
- ^ Sheppard, K. A.; Rival, D. E.; Caron, J.-B.. “On the Hydrodynamics of Anomalocaris Tail Fins” (英語). Integrative and Comparative Biology. doi:10.1093/icb/icy014.
- ^ a b c De Vivo, Giacinto; Lautenschlager, Stephan; Vinther, Jakob (2016-12-16). Reconstructing anomalocaridid feeding appendage dexterity sheds light on radiodontan ecology.
- ^ Paterson, John R.; García-Bellido, Diego C.; Lee, Michael S. Y.; Brock, Glenn A.; Jago, James B.; Edgecombe, Gregory D. (2011-12). “Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes” (英語). Nature 480 (7376): 237–240. doi:10.1038/nature10689. ISSN 0028-0836.
- ^ Jay,, Gould, Stephen. Wonderful life : the Burgess Shale and the nature of history (First edition ed.). New York. ISBN 0393027058. OCLC 18983518
- ^ a b Whittington, Harry Blackmore; Briggs, Derek Ernest Gilmor (1985-05-14). “The largest Cambrian animal, Anomalocaris, Burgess Shale, British-Columbia” (英語). Phil. Trans. R. Soc. Lond. B 309 (1141): 569–609. doi:10.1098/rstb.1985.0096. ISSN 0080-4622.
- ^ a b c Daley, Allison; Bergström, Jan (2012-04-05). “The oral cone of Anomalocaris is not a classic ‘‘peytoia’’”. Die Naturwissenschaften 99: 501–4. doi:10.1007/s00114-012-0910-8.
- ^ Van Roy, Peter; Briggs, Derek E. G. (2011-05). “A giant Ordovician anomalocaridid” (英語). Nature 473 (7348): 510–513. doi:10.1038/nature09920. ISSN 0028-0836.
- ^ Smith, Martin R.; Ortega-Hernández, Javier (2014-08-17). “Hallucigenia’s onychophoran-like claws and the case for Tactopoda”. Nature 514 (7522): 363–366. doi:10.1038/nature13576. ISSN 0028-0836.
- ^ Briggs, Derek E. G. (2015-04-19). “Extraordinary fossils reveal the nature of Cambrian life: a commentary on Whittington (1975) ‘The enigmatic animal Opabinia regalis, Middle Cambrian, Burgess Shale, British Columbia’” (英語). Phil. Trans. R. Soc. B 370 (1666): 20140313. doi:10.1098/rstb.2014.0313. ISSN 0962-8436. PMID 25750235.
- ^ Haug, Joachim; Waloszek, Dieter; Maas, Andreas; Liu, Yu; Haug, Carolin (2012-03-01). “Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian”. Palaeontology 55: 369–399. doi:10.1111/j.1475-4983.2011.01124.x.
- ^ Park, Tae-Yoon S.; Kihm, Ji-Hoon; Woo, Jusun; Park, Changkun; Lee, Won Young; Smith, M. Paul; Harper, David A. T.; Young, Fletcher et al. (2018-03-09). “Brain and eyes of Kerygmachela reveal protocerebral ancestry of the panarthropod head” (英語). Nature Communications 9 (1). doi:10.1038/s41467-018-03464-w. ISSN 2041-1723.
- ^ 土屋健『エディアカラ紀・カンブリア紀の生物』(技術評論社 2013)
- ^ “Comment on “Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris” by Stephen Pates, Allison C. Daley, and Javier Ortega-Hernández - Acta Palaeontologica Polonica” (英語). www.app.pan.pl. 2018年9月19日閲覧。
- ^ Jan Bergström & Hou Xian-Guang (2003). “Arthropod origins” (PDF). Bulletin of Geosciences 78 (4): 323–334.
- ^ Caron, J. -B.; Gaines, R. R.; Mangano, M. G.; Streng, M.; Daley, A. C. (2010). “A new Burgess Shale-type assemblage from the "thin" Stephen Formation of the southern Canadian Rockies”. Geology 38 (9): 811. doi:10.1130/G31080.1.
- ^ Yuanyuan, Wang; Huang, Diying; Hu, Shixue (2013-11-01). “New anomalocardid frontal appendages from the Guanshan biota, eastern Yunnan”. Chinese Science Bulletin 58. doi:10.1007/s11434-013-5908-x.
- ^ a b Vinther, J.; Stein, M.; Longrich, N. R.; Harper, D. A. T. (2014). “A suspension-feeding anomalocarid from the Early Cambrian”. Nature 507: 496–499. doi:10.1038/nature13010. PMID 24670770.
- ^ Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A.; Shu, Degan; Paterson, John R.. “Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan” (英語). National Science Review. doi:10.1093/nsr/nwy057.
- ^ Daley, Allison C.; Budd, Graham E.; Caron, Jean-Bernard; Edgecombe, Gregory D.; Collins, Desmond (2009). “The Burgess Shale anomalocaridid Hurdia and its significance for early euarthropod evolution”. Science 323 (5921): 1597–1600. doi:10.1126/science.1169514. PMID 19299617.
- ^ STEIN, MARTIN (2010-02-26). “A new arthropod from the Early Cambrian of North Greenland, with a ‘great appendage’-like antennula” (英語). Zoological Journal of the Linnean Society 158 (3): 477–500. doi:10.1111/j.1096-3642.2009.00562.x. ISSN 0024-4082.
- ^ Legg, David (2013-05-01). “Multi-Segmented Arthropods from the Middle Cambrian of British Columbia (Canada)”. Journal of Paleontology 87: 493–501. doi:10.1666/12-112.1.
- ^ The Cambrian fossils of Chengjiang, China : the flowering of early animal life. Hou, Xianguang. (Second edition ed.). Chichester, West Sussex. ISBN 9781118896310. OCLC 970396735
- ^ a b Xianguang, Hou; Bergström, Jan; Jie, Yang (2006). “Distinguishing anomalocaridids from arthropods and priapulids” (ドイツ語). Geological Journal 41 (3-4): 259–269. doi:10.1002/gj.1050. ISSN 1099-1034.
- ^ Chen, Jun-yuan; Ramsköld, Lars; Zhou, Gui-qing (1994-05-27). “Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators” (英語). Science 264 (5163): 1304–1308. doi:10.1126/science.264.5163.1304. ISSN 0036-8075. PMID 17780848.
- ^ “Anomalocaridids and the origin of arthropods: the view from Chengjiang” (英語). ResearchGate. 2019年1月19日閲覧。
参考文献
- Briggs, Derek; Collier, Frederick; Erwin, Douglas. The Fossils of the Burgess Shale. Smithsonian Books, 1995.
- James W. Valentine. On the Origin of Phyla. University Of Chicago Press, 2004.
- Tim Haines & Paul Chambers. The Complete Guide to Prehistoric Life. BBC Books, 2005.
- Conway Morris, Simon. The Crucible of Creation. Oxford University Press, 1998.