
鋏角類
| 鋏角類 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 様々な鋏角類[注釈 1]
| |||||||||
| 地質時代 | |||||||||
| 古生代カンブリア紀 - 現世 | |||||||||
| 分類 | |||||||||
| |||||||||
| 学名 | |||||||||
| Chelicerata Heymons, 1901 | |||||||||
| 英名 | |||||||||
| Chelicerate | |||||||||
| 下位分類群 | |||||||||
鋏角類(きょうかくるい、英語: Chelicerate、学名: Chelicerata[2])は、節足動物を大まかに分ける分類群のひとつである。分類学上では鋏角亜門とされ、クモ・サソリ・ダニなどのクモガタ類・およびカブトガニ・ウミサソリ・ウミグモなどを含む。1対の鋏角と、複数対の歩脚型の肢を体の前方にもつ[3]。
11万種以上が知られ[4][5]、現生節足動物の4つの亜門の中では2番目に種を富んだ分類群である。系統関係については、多足類・甲殻類・六脚類という3つの亜門を含んだ大顎類と姉妹群になり、現生節足動物を大きく分けた2つの系統群の一角になる[5][6]。
学名「Chelicerata」およびその由来になった鋏角の英語名「chelicera」はギリシア語の「khēlē」(鋏)と「keras」(角)の合成語である[7][8]。
形態
[編集]
![サソリの構造 各項説明:[注釈 2]](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Scorpion_Tagged_(PSF).png)


鋏角類に含まれる節足動物は次の基本的体制の違いによって、それ以外の節足動物(大顎類・三葉虫など)から区別できる。
- 体の合体節(tagma)は原則として前後で前体と後体の2部のみである[注釈 3][3]。
- 第1体節由来の付属肢は鋏角である[3](大顎類と三葉虫の場合は触角である)[9]。
- 先頭の合体節(頭部融合節)における鋏角以外の付属肢は原則として歩脚型である[3](大顎類の場合は触角と顎に特化している)[9]。
- 触角と顎は存在しない(似た機能に収斂した相似の付属肢もしくはその一部をもつ例のみ存在する[注釈 4])[3][10]。
前体
[編集]



前体(prosoma)は眼と口をもつ先節と直後の第1-6体節からなる合体節である。通常、前体は全ての体節が融合して背面は1枚の背甲(carapace、甲殻類の背甲から区別するために prosomal dorsal shield や peltidium とも呼ぶ[11][3])に覆われるが、第5と第6体節が独自に分節した例も存在する[注釈 5][3][12]。腹面中央に配置される腹板(sternum、または胸板[13]、節口類の場合は endostoma)は分類群によってあったり欠けたりする[3]。大顎類に見当たる触角と顎は存在しないが、それらに類する感覚用と摂食用の相似器官をもつ例が多く挙げられる[注釈 4][10]。
他の節足動物の頭胸部との違い
[編集]この部分は「頭胸部」(cephathorax)とも呼ばれていたが、鋏角類の前体は1つの合体節として認められ、体節制や遺伝子発現的にも頭部そのものに該当する頭部融合節であり[14][15][16][17][18][19]、他の節足動物(例えば甲殻類)の頭胸部のように頭部と胸部という2つの合体節を含んだ部分ではない[9]。節足動物全般の頭部構成に関する議論、特に他の節足動物の頭部と比較する場合、鋏角類の前体は常に「頭部」扱いとされる[14][20][21]。すなわち、大顎類において顎に特化した付属肢は、鋏角類の場合ではそのほとんどが脚として用いられ[22]、「頭で歩いている」とも比喩される[3](後述の対応関係および節足動物#系統関係と体節の相同性も参照)。
眼
[編集]鋏角類の前体背面は、原則として左右に側眼(lateral eye)、中央に中眼(median eye)を有し、それぞれ祖先形質として複眼と単眼である[23][24]。ウミグモの場合は側眼はなく、原則として2対中眼のみをもつ[23]。それ以外の鋏角類の中眼は通常1対[注釈 6]で、カブトガニ類とウミサソリ類の側眼はれっきとした複眼である。基盤的なサソリ類と一部のワレイタムシもそれに近い側眼をもつ[23][25]が、現生のクモガタ類はそれが退化し、複眼の個眼に由来する数対以下の単眼となる[23][24][26]。なお、コヨリムシのように、全ての眼を退化消失した鋏角類もある[23][26]。
前体の付属肢
[編集]
I:鋏角
II:触肢
III-VI:脚


体節数に応じて、前体は原則として6対の付属肢(関節肢)がある[3]。第1体節は本群の最も重要な共有派生形質[27][5][3]である1対の鋏角(きょうかく、chelicera, 複数形: chelicerae, ウミグモの場合は鋏肢 chelifore という)をもつ。通常は目立たない鋏型の付属肢であるが、分類群によっては巨大化したり[注釈 7]牙のような形[注釈 8]になったりする場合もある[3]。鋏角は原則として3節の肢節(第1肢節は柄部、第2肢節は掌部と不動指、第3肢節は可動指[3])に分かれるが、柄部を欠けて2節になる分類群はクモガタ類に多く見られ[注釈 9]、イトダニ科のダニと一部のウミグモ類はその柄部が2節以上に分かれ、計4節以上に分かれた鋏角をもつ[21][3][28]。
次の第2-6体節は、7節前後の肢節に分かれた5対の歩脚型付属肢がある[3]。そのうち最初の1対は触肢(しょくし、pedipalp、ウミグモの場合は palp)といい、クモガタ類とウミグモ類の場合ではこの付属肢の特化が進んでおり、明確に脚から区別される[注釈 10][3]。この5対の付属肢はほとんどの場合は内肢のみをもつ単枝型であるが、ごく一部の化石群においては外肢が見られる二叉型で、特にオファコルスやダイバステリウムなど基盤的な群では、最初の4対は内肢に劣らないほど発達した外肢をもつ[29][30]。これらの化石種の特徴に基づいて、鋏角類のこの5対の付属肢はかつて外肢があり、現生群に至る系統でそれがほぼ完全に退化消失していたと考えられる[注釈 11][30][31][3][1]。
通常、鋏角類の前体付属肢は全てが機能的であるが、そのいずれが二次的に退化消失した例もごく稀にある[注釈 12][3][32]。
鋏角はクモの場合では「上顎」とも呼ばれ、触肢や脚の基部に備わる突起物は分類群によって「顎基」「顎葉」「下顎」などと呼ばれるが、いずれも大顎類の節足動物の顎(大顎と小顎)とは別起源で、機能が相似するに過ぎない[10]。
口と周辺の構造
[編集]口は鋏角と触肢の間に開き、目立たない上唇(labrum、または rostrum[11])に覆われている。カブトガニ類の口は後方に向くが、クモガタ類の場合は前方に向く[3]。ウミグモ類の場合は上唇らしき構造をもたず、代わりに発達した円筒状の吻(proboscis)が先頭に突出し、口はその先端に開く[3]。
後体
[編集]
後体(opisthosoma)は腹部(abdomen)とも呼ばれるが、合体節的には胴部である。第7体節を起点として、最多13節からなる(第19体節まで及ぶ)[3]。通常は単一の合体節とされるが、前後で幅広い前部と細い後部に特化し、いわゆる中体(mesosoma)と終体(metasoma)もしくは前腹部(preabdomen)と後腹部(postabdomen)という、さらに2つの合体節として明瞭に区別できた分類群もある[注釈 13]。また、クモガタ類の中には尾部(pygidium)と呼ばれる、短く集約した末端2-3節をもつ分類群もある[注釈 14]。

最終体節の肛門の直後に尾節(telson)をもつ例が多く、その形は分類群によって剣状(棘状)・へら状・鞭状(数珠状、鞭状体 flagellum)など様々である[注釈 15][3]。
一部の文献では「中体・後体」と「前腹部・後腹部」は違う定義で区別され、中体と後体は(特化の有無にかかわらず)それぞれ後体第1-7節(第7-13体節)と後体第8節(第14体節)以降の体節を専門に示し、前腹部と後腹部はそれぞれ単に後体の幅広い前部と細い後部を示す用語(体節の番目にこだわらないため、それが必ずしも前述の中体と後体の範囲に対応するとは限らない)として用いられていた[31][3]。しかし、これは後述の後体付属肢の配置や「中体と終体に特化した後体は真鋏角類の祖先形質」という仮説に基づいた区別方法であり[31]、全ての文献に採用されるとは限らない[注釈 16]。
後体の付属肢
[編集]
-

-
 糸を出している最中のクモの糸疣
糸を出している最中のクモの糸疣 -




通常、後体の付属肢はほとんどが退化的で、あっても原則として後体第1-7節(第7-13体節)の範囲内のみに生えて[注釈 17]、前体の付属肢からかけ離れた形態をもつ。節口類と蛛肺類[注釈 18]の生殖器や呼吸器(書鰓と書肺)を腹面から覆いかぶさった板状構造体は蓋板(がいばん、operculum, 複数形: opercula, 鰓蓋(えらぶた)とも)という付属肢であり[33][3]・クモの糸疣・サソリの櫛状板・ウミサソリ、ウデムシとサソリモドキの生殖肢などの器官も、付属肢由来の器官だと考えられる[34][33][3][35]。また、蓋板をもつ体節は腹板が退化的な場合が多く、これは四肺類[注釈 8]で特に進んでおり、元の腹板は外から観察できず、代わりに蓋板がまるで腹板のように体と密着していた[36][37][33]。
第7体節
[編集]
後体の中で、前体との境目に当たる最初の1節、いわゆる「第7体節」はその性質によって後体的本質が疑問視される場合がある[3][38]。クモガタ類の場合、この体節は付属肢をもたず[注釈 19][3]、一部の群ではくびれて腹柄(pedicle)に特化した[3]が、その腹板が前体の範囲に食い込んだ例も見られる[36][37]。カブトガニ類の場合、この体節は前体と融合して背甲に覆われ、1対の唇様肢(chilaria)という前体の付属肢とセットに機能した付属肢をもつ[3][31]。ウミサソリ類もそれに似て、左右融合した第7体節付属肢と思われる1枚の下層板(metastoma)を有し、前体に食い込むように最終の脚の間に配置される[3]。ウミグモ類の場合はさらに異様で、この体節は前体と同形の脚が生えて[3]、ハラフシカブトガニ類のウェインベルギナもこの特徴をもつかもしれない[3]。基盤的な鋏角類とされる化石節足動物(ハベリア、モリソニアなど)のこの体節も、前体の一部として機能する傾向がある[38][1]。これらの特徴を基に、第7体節の多くの性質は前体的で、むしろ前体の一部と扱うべきではないかという提唱もある[3][31]。一方、ホメオティック遺伝子発現では、この体節は前体と後体の境目的である[16][17][18][19]。
呼吸器
[編集]
鋏角類は全般的に多様な呼吸様式が見られる[3]。水生鋏角類であるカブトガニ類とウミサソリ類の後5対の蓋板は、後側に書鰓(しょさい、book gills)という本のページのように畳んだラメラ(lamellae、薄葉、薄板)で構成される呼吸器をもつ[3][39]。ウミサソリの場合はさらに特化が進み、「kiemenplatten」という本群に特有の呼吸器は蓋板と書鰓にあわせて精密な鰓室(gill chamber)を構成し[3]、その書鰓から空気呼吸に適した構造体も発見される[39]。クモガタ類の中で蛛肺類[注釈 18]は書鰓由来と思われる書肺(しょはい、book lungs)をもち[3]、それ以外の群は多くが気管(trachea)で[注釈 20][3]、コヨリムシ、一部のダニ、およびウミグモ類は皮膚呼吸のみを通じて酸素を取り込む[40]。小型の鋏角類ほど、呼吸の様式は書肺より気管や皮膚呼吸に偏って単純化する傾向がある[32]。
書鰓と書肺は蓋板の器官であるため後体のみに備わるが、気管は分類群によって後体のみ(ザトウムシ、カニムシ、アシナガダニ)、前体のみ(クツコムシ、一部のダニ)、もしくは後体と前体両方(ヒヨケムシ)に備わるものがある[3]。書肺と気管の開口部は気門(spiracle)といい、一部の例外[注釈 21]を除いてこれは常に体の腹面で対になって開口する[3]。
生殖器
[編集]_Figures_14%E2%80%9318.jpg)
-
-

-
 クモのオスの触肢器
クモのオスの触肢器 -


.jpg)

_Figure_8.jpg)
多くの場合、鋏角類はペニスや産卵管と言えるほど顕著な外性器はなく、生殖器の外部は生殖孔(gonopore)のみによって表れる。ウミグモ類の生殖孔は各脚の基部に開いている[41]が、それ以外の鋏角類では知られる限り後体第2節(第8体節)の腹側のみに開口し[11][3]、該当体節の蓋板もしくは腹板由来の生殖口蓋(genital operculum)に覆われる[3]。生殖孔に当たる部分で外性器をもつ例として、ウデムシとサソリモドキは生殖口蓋の裏側に1対の小さな生殖肢(gonopod)[3]、ウミサソリ類とカスマタスピス類は生殖口蓋の中央に「genital appendage」という棒状の生殖肢をもつ[42][43]。ザトウムシは例外的に雌雄それぞれ発達したペニスと産卵管をもち[3]、クツコムシのオスも生殖孔にペニスと呼ばれるほど突出した構造体がある[44]。生殖孔でない部分にあるものの配偶行動に直結する器官は、クモの触肢にある触肢器(palpal organ, 移精器官)と、ヒヨケムシのオスの鋏角にある鞭毛(flagellum)が挙げられており、クツコムシのオスの第3脚にも「copulatory organs」という同じ機能をもつと思われる器官がある[3]。
神経系
[編集]
各項説明:[注釈 22]
-
 ウミグモ類の神経系(黄、B)
ウミグモ類の神経系(黄、B) -
 カブトガニ類の神経系
カブトガニ類の神経系 -
![ウデムシの神経系 各項説明:[注釈 23]](//upload.wikimedia.org/wikipedia/commons/thumb/9/9e/Phrynus.cns.boerner.1904.png/217px-Phrynus.cns.boerner.1904.png)
-
 クモの神経系(青)
クモの神経系(青)

_(7362373642).jpg)
![ウデムシの神経系 各項説明:[注釈 23]](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Phrynus.cns.boerner.1904.png)

他の節足動物と同様、鋏角類ははしご形神経系をもつ。脳(brain もしくは syncerebrum[45])に含まれる神経節の体節や付属肢との対応関係はかつて議論があったが(後述参照)、2010年代以降では先節の神経節は前大脳(protocerebrum)、第1体節/鋏角の神経節は中大脳(deutocerebrum)、第2体節/触肢の神経節は後大脳(tritocerebrum)という解釈が確定的で、それ以降の体節/付属肢の神経節は腹神経索(ventral nerve cord)と扱うのが一般的である[20]。ウミグモ類の脚の神経節ははっきりとしたはしご形を残るが、カブトガニ類とクモガタ類の場合、脳神経節と脚の神経節は融合が進み、脳と腹神経索の区分がほぼなくなり、前体全ての神経節が「synganglion」という1つの集中部になっている[45]。食道はその中央(神経解剖学的には中大脳の間[46])を貫通し、これに基づいて synganglion を前後で食道上神経塊(supraesophageal ganglion)と食道下神経塊(subesophageal ganglion)として区別される場合もある[47]。後体の神経節の場合、カブトガニ類とサソリ類ははしご形のままであるが、他のクモガタ類は多くが前体に集約される[45]。側眼の視神経が3つの神経網をもつ大顎類とは異なり、現生鋏角類の側眼の視神経は1つの神経網のみをもつ[24]。
大きさ
[編集]
-
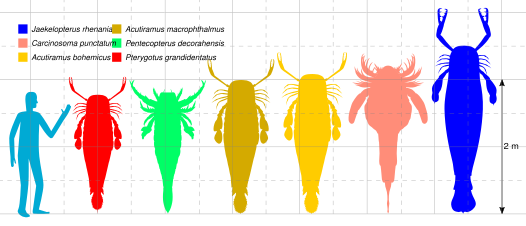 最大級のウミサソリ類の比較図
最大級のウミサソリ類の比較図

鋏角類の大きさは多様で、現生群だけでも数十cm程度のカブトガニ類から、80–200μm程度の小型ダニ類まで挙げられる[32]。様々な分類群(目)の中で、ダニは最も小型化が進み、体長が1mmも満たさない種類が多い[32]。クモは体格差が最も極端で、知られる中で最大(約10cm)と最小(0.37mm)の種類は270倍の体長差にある[32]。化石群まで範囲を広げると、ウミサソリ類は1m前後の大型種を数多く含み[49]、中で2.5m程度の巨体をもつと推測され、知られる中で最大級の節足動物として知られるものもある[50]。
生態
[編集]生息地
[編集]




鋏角類の中で、クモガタ類はほとんどが陸生で様々な陸上生態系に進出し、砂漠・極地・高山など極端な環境に生息する種類もある[51]。水中生態系の場合、クモガタ類にはミズダニやミズグモなど陸から二次的に水生化したものが知られ[51]、カブトガニ類とウミグモ類は現生群ではすべてが海棲である。しかし化石群まで範囲を広げると、カブトガニ類とウミサソリ類はいずれも海棲・陸水性と考えられる種類を数多く含み、特にウミサソリ類は、前述の特殊な呼吸器によって陸上でも呼吸でき[39]、少なくともある程度の陸上活動をできたと思われる種類もある[52][49][39]。
食性
[編集]
-
 草食性のハダニ。
草食性のハダニ。 -

-


.jpg)

鋏角類は一般に捕食者のニッチ(生態的地位)を占め、特にクモガタ類は多くが昆虫などの小型節足動物を捕食する肉食動物で、絶滅群のウミサソリ類も多くが獰猛な捕食者であったと考えられる。しかし、幾つかの例外も挙げられる。例えばクモガタ類の中でダニ類は例外的で肉食性・草食性・腐植食性・寄生性・吸血性まで多岐にわたり[53]、ザトウムシ類は雑食性や腐肉食性の種類によって知られる。捕食者として代表的であるクモ類の中でも吸蜜行動が見られ[54][55]、植物成分を主食とする1種も報告される[56]。現生のカブトガニ類は肉食性に偏る雑食性である[57]。ウミグモ類の食性については研究が進んでいないが、主に刺胞動物など柔らかい固着生物の体内組織を摂ることが知られ、これは文献によって捕食性もしくは寄生性と扱われる[58]。

多くの場合、鋏角類の鋏角は口器として機能する主な部分であり、餌を把握・切断・粉砕するのに用いられる。クモ類の場合、牙のような鋏角は毒腺をもち、獲物を麻痺させる機能も兼ね備える[3]。鋏角以外の前体付属肢(触肢や脚)の基部が摂食機能を担う構造をもつ例も多く、クモ類の下顎(maxilla、触肢の基部に備わる)・ザトウムシ類とサソリ類の顎葉(coxapophyses、ザトウムシの場合は触肢と第1脚の基部、サソリの場合は第1-2脚の基部に備わる)[59]・カブトガニ類とウミサソリ類の顎基(gnathobase、触肢と全ての脚の基部に備わる)などが挙げられる[10]。カブトガニ類とウミサソリ類のそれぞれの後脚の間にある唇様肢と下層板も、口器として機能する付属肢とされる[10]。ウミグモ類は突き出した吻で直接に餌を摂るが、発達した鋏肢と触肢をもつ種類ではこれらの付属肢も摂食を補助する役割を果たしている[58]。
多くのクモガタ類は固形物の餌を直接に摂食せず、代わりに消化液で餌を体外消化し、軟組織を液体状に分解してから口内に飲み込むが、ザトウムシ類は例外的に固形物を直接に摂食できる[5]。現生のカブトガニ類は発達した前胃で固形物の餌を細かく砕き、食べられない物をここから噴き返すこともできる[60]。
繁殖と発育
[編集]
_(8390306848).jpg)


鋏角類の中で様々な繁殖行動が見られ、クモガタ類の中では特殊な求愛行動をもつ分類群もいくつかある。配偶子のやりとりとして、ザトウムシ類は交尾(雌雄生殖器の連結を通じて行う)、他のクモガタ類は交接(精包の受け渡しなど、交尾以外のあらゆる方法で精子をメスの生殖孔に入り込む)、カブトガニ類とウミグモ類は体外受精を通じて行う[41]。原則として卵生だが、サソリは卵胎生である。卵や幼生の世話をする保育行動は、クモガタ類とウミグモ類で普遍に見られる[41]。
他の節足動物と同様、鋏角類は脱皮で成長し、幼生は多くの場合では成体と同じ体節数と付属肢数で生まれる。例外として、生まれたての多くのダニ類とクツコムシ類は6本の脚のみをもち、後に脱皮してから8本脚となる。ウミグモ類はプロトニンフォン幼生(protonymphon)という成体らしからぬ特殊な形態で生まれ、脱皮を通じて徐々に脚をもつ体節を増やして成体に近い姿に変態する[61][62]。また、性成熟を迎えると成長が止まるもの(カブトガニ類、多くのクモガタ類など)と、性成熟になっても脱皮し続けられるものがある(オオツチグモのメス、ウデムシなど)[63]。欠損した付属肢は通常では次の脱皮である程度まで再生できるが、付属肢の再生能力を欠く分類群もある(ザトウムシなど)[64]。
体節と付属肢の対応関係
[編集]
| 分類/体節(先節を除く) | 1(中大脳) | 2(後大脳) | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Artiopoda類 | 触角 | 第1脚 | 第2脚 | 第3脚 | 第4脚 | 第5脚 |
| 大顎類 (多足類・甲殻類・六脚類) |
第1触角 | 第2触角/(退化) | 大顎 | 第1小顎 | 第2小顎/下唇 | 第1脚 |
| 鋏角類 (鋏角-大顎相同説) |
(退化) | (退化) | 鋏角 | 触肢 | 第1脚 | 第2脚 |
| 鋏角類 (20世紀の主流な見解) |
(退化) | 鋏角 | 触肢 | 第1脚 | 第2脚 | 第3脚 |
| 鋏角類 (21世紀以降の見解) |
鋏角 | 触肢 | 第1脚 | 第2脚 | 第3脚 | 第4脚 |
鋏角類の体制は他の節足動物と大きく異なるため、外見から他の節足動物との付属肢や体節の対応関係(相同性)は判断しにくく、特に触角を持たない原因や鋏角の由来が議論の的となった。研究史上では複数の相容れない対立仮説を提唱され、次の通りに挙げられる[8]:
- 大顎類のどの付属肢にも相同でない[66]。
- 第3体節由来:大顎類の大顎に相同とされる[67]。
- 後大脳性/第2体節由来:甲殻類の第2触角に相同(多足類と六脚類の場合は付属肢をもたない間挿体節)とされる[68]。
- 中大脳性/第1体節由来:大顎類の触角(甲殻類の第1触角)に相同とされる[68][69]。
いずれも19世紀から既に提唱された説である[66][68][67][69]が、当時では「鋏角類」という分類群は未創設で(後述参照)、クモガタ類と節口類の鋏角は別々に大顎(クモガタ類)と触角(節口類)に相同ともされていた[70][8]。クモガタ類と節口類をまとめる「鋏角類」が創設された20世紀では、鋏角の大顎との相同性は否定され、後大脳性/第2体節由来説が主流となり、同じ時期で主流になった鋏角類と三葉虫など(Artiopoda類)の類縁関係も、この説を踏まえて議論をなされていた(後述参照)[71][72]。
しかし21世紀以降では、次のホメオボックス遺伝子発現・発生学・神経解剖学で得られる情報により、中大脳性/第1体節由来説と第3体節由来説は徐々に否定され、後大脳性/第2体節由来説が確定的になった。
2000年代では、同じ鋏角類の中で、ウミグモ類と他の鋏角類の体節/付属肢の対応関係が疑問視されることも一時的にあったが、否定的とされる(詳細はウミグモ綱#頭部付属肢の対応関係を参照)[78][75]。
-
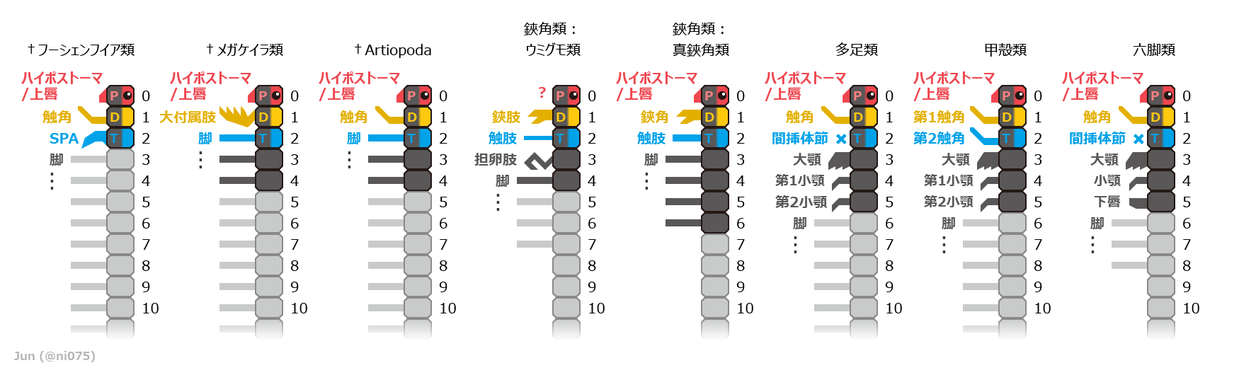 2017年時点で広く認められる、節足動物の主要な高次系統における付属肢と体節の対応関係。
2017年時点で広く認められる、節足動物の主要な高次系統における付属肢と体節の対応関係。

系統関係
[編集]
| 節足動物門 |
| ||||||||||||||||||




20世紀以前では、節足動物の中で「鋏角類」という分類群はなく、クモガタ類は昆虫と何らかの繋がりをもつとされ[65]、節口類は甲殻類に分類されていた[70]。しかし20世紀初期の再検討をはじめとして、クモガタ類と節口類の体制は昆虫や甲殻類に似ておらず、むしろお互いに独自の体制を共有することが徐々に明らかになった[65]。こうして1901年、ウミグモ類・節口類・クモガタ類をまとめた節足動物の分類群「鋏角類」(Chelicerata)が創設された[2]。
現生節足動物の4つの亜門の中で、鋏角類は最初に分岐した単系統群で、残り全ての現生節足動物(大顎類 Mandibulata)の姉妹群になるという系統位置は、ホメオティック遺伝子発現[15]、および多くの形態学と分子系統学的見解に強く支持される[79][6]。他にも甲殻類などと姉妹群になる(Schizoramia をなす)、多足類と単系統群になる(多足鋏角類 Myriochelata /矛盾脚類 Paradoxopoda をなす)、もしくは鋏角類は多系統群(幹性類 (Cormogonida) 説、後述参照)などの異説はかつてあったが、いずれも後に否定的とされる[6]。
上述の現生群以外にも、鋏角類と化石節足動物の分類群の類縁関係は多くの議論が繰り広げられ、次の通りに挙げられる。
化石節足動物との関係性
[編集]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 現生鋏角類の分類群(太字)といくつかの化石節足動物(†)の系統関係。姉妹群関係が不確実のものは複数分岐としてまとめられる。 |


_003.jpeg)



.png)

鋏角類はおよそ5億年前の古生代カンブリア紀に起源と考えられ[1]、基盤的な鋏角類とされるカンブリア紀の化石節足動物は、主にメガケイラ類(Megacheira, 大付属肢節足動物)、ハベリア類(ハベリア目 Habeliida)、およびモリソニア類(モリソニア目 Mollisoniida)が挙げられる[80]。メガケイラ類は鋏角類に似た中枢神経系[81]、退化的な上唇[82]、および鋏角を彷彿とさせる構造の大付属肢[83]があり、後述の群ほど確定的ではないが、これらの性質で類縁関係を示唆される[84][83][85][86][5][87]。サンクタカリスやハベリアなどを含んだハベリア類は、再検討により鋏角類の体制をもつことが判明し、前体の付属肢には基盤的な真鋏角類であるオファコルスやダイバステリウムに似た外肢もある[88][38]。しかし、これらの節足動物は鋏角に相同と思われる付属肢をもつものの、そのいずれも確実に鋏角といえるほどの構造ではなかった[1]。モリソニア類のモリソニア[89]は、真の鋏角をもつことが認められる初のカンブリア紀節足動物で、真鋏角類的な中枢神経系と書鰓を彷彿とさせる構造も兼ね備えていた[1][87]。
-
 様々なメガケイラ類
様々なメガケイラ類 -

-


それ以外の化石節足動物との系統関係については、三葉虫類(Trilobita)などの節足動物を含んた分類群三葉形類(Trilobitomorpha)に類縁で、Arachnomorphaを構成する説は、20世紀においては主流であった[34][5]、これは主にカブトガニ類と三葉形類の類似点(幅広い背甲・後体/胸部の三葉構造・平板状の蓋板/外肢・顎基をもつ前体/頭部付属肢など)が根拠とされ[72][71][84][85][5]、その中で鋏角類は三葉形類から派生するという考えもあった[71][72]。同時に光楯類(Aglaspidida)は、鋏角類のように頭部(前体)に鋏角を含めて6対の付属肢をもつと解釈され、節口類の鋏角類と考えられた[90][31]。また、三葉形類は触角をもつため、鋏角類は三葉形類らしき共通祖先から派生する過程で、触角(およびそれをもつ第1体節)を二次的に退化したと考えられ[34][72][84]、ケロニエロンなど一部の化石節足動物が、その中間型生物ともされてきた[71]。
しかし、20世紀後期の研究を始めとして、光楯類は鋏角類でない(頭部の付属肢は4-5対しかなく、最初の付属肢は鋏角ではなく触角である)ことが徐々に判明し(光楯類#分類を参照)、90年代以降では鋏角類の鋏角は第1体節由来で、他の節足動物の第1触角に相同であることも分かり(前述参照)、三葉形類との類似点もカブトガニ類以外の鋏角類に対応できず、類縁関係を示唆する根拠として疑わしく見受けられるようになった[91]。これにより、前述の一連の系統仮説はほぼ無効化され、新たな基準で見直さなければならない[85]。
21世紀以降では、三葉形類と光楯類はまとめてArtiopoda類に分類されるようになり[90]、その中で根拠は20世紀と多少異なりながら、Arachnomorpha説(鋏角類+Artiopoda類)を支持する研究はいくつかある[92][88][38]。しかしこれは確実でなく、代わりにArtiopoda類と大顎類の類縁関係(Antennulata説)を支持する研究も少なからぬ挙げられる[91][5][93][6]。
| 節足動物 |
| ||||||||||||
| 節足動物 |
| ||||||||||||
節足動物と考えられるラディオドンタ類(Radiodonta、アノマロカリスなどを含む群)は、メガケイラ類と鋏角類のように、口の前に第1触角の代わりに捕獲用の附属肢(前部付属肢)があるため、両者に類縁と考えられることもあった[94][86]。そこで、初期の節足動物の中で鋏角類に至る系統群はまず前端の付属肢を捕獲用に特化し、ラディオドンタ類の前部付属肢はメガケイラ類の大付属肢に、大付属肢は鋏角類の鋏角に進化するという説が提唱された[83][95][86]。しかし、ラディオドンタ類の多くの祖先形質や神経解剖学的証拠はこのような系統仮説を支持しなかった[20]。メガケイラ類と鋏角類は真節足動物であり、それぞれの大付属肢と鋏角は、他の節足動物の第1触角と同様に中大脳性(第1体節由来)である[81][80]のに対して、ラディオドンタ類は真節足動物より早期に分岐した基盤的な節足動物として広く認められ[96][20]、その前部付属肢も前大脳性(先節由来)で、別起源の可能性が示される[97]。
下位分類
[編集]-
 ウミグモ類
ウミグモ類 -
 広義のカブトガニ類(1-4:狭義のカブトガニ類、5-6:ハラフシカブトガニ類)
広義のカブトガニ類(1-4:狭義のカブトガニ類、5-6:ハラフシカブトガニ類) -

-

-

)_(20928914062).jpg)



基盤的な鋏角類とされる前述の古生物を除き、通常、鋏角類は大きくウミグモ類(Pycnogonida、ウミグモ綱、皆脚綱)と真鋏角類(Euchelicerata)の2群に分かれ、後者はさらにカブトガニ類・ウミサソリ類などを含んだ節口類(Merostomata、節口綱、腿口綱)、およびクモ・サソリ・ダニなどの陸生群を含んだクモガタ類(Arachnida、クモガタ綱、蛛形綱、クモ綱)として細分される[5]。
しかしこのような分類体系の一部は単系統性を反映していないと疑問視され、以下の議論が繰り広げられる。
| |||||||||||||||||||||||||||
| 一般的な鋏角類の系統仮説。(青桁=節口類) |
| |||||||||||||||||||||
| 真鋏角類の中で、クモガタ類がカブトガニ類とウミサソリ類に対して非単系統である系統仮説。(青桁=節口類) |
| |||||||||||||||
| 幹性類説。ウミグモ類の系統位置によって鋏角類(*)は解体される。 |
- クモガタ類:
- 四肺類(クモ・ウデムシ・サソリモドキ・ヤイトムシなどを含んだ群)の単系統性は昔今を通じて広く認められ[5]、かつて別系統と考えられたサソリとの姉妹群関係(蛛肺類)も後に様々な情報(分子系統学[79][98][99][100]・遺伝子重複[101][102]・書肺の連続相同性[103])で支持が得られる[51]。しかしそれ以外の群の類縁関係は、主に分子系統解析によって不確定の状態に至った[5][6]。ダニの単系統性は賛否両論で[98][104][100]、またクモガタ類自体が他の鋏角類(ウミグモ類[105]・ウミサソリ類[51]・カブトガニ類[106][98][99][107][108]のそのいずれ)に対して非単系統ではないかと思われることもしばしばある[5][109]。→詳細は「クモ綱 § 分類と進化」を参照
- 節口類:
- 節口類は一般にクモガタ類より早期に分岐した群とされるが、前述のクモガタ類非単系統説により、特定のクモガタ類(特に蛛肺類)に近縁とされる場合もある[51][99][107][39]。化石群であるウミサソリ類は、主に生殖器官の形質に基づいてカブトガニ類よりもクモガタ類に近縁であるとされる[110][111][112][31]。こうして節口類はクモガタ類に対して側系統群である説が一般的になり、「節口類」を廃止させる見方もある[31]。また、従来ではカブトガニ類の一部としてまとめられた化石群ハラフシカブトガニ類[113](Synziphosurina)は、他のカブトガニ類(狭義のカブトガニ類)に類縁せず、むしろ他の真鋏角類全般に対して非単系統である説が後に有力視される[31][114][115][116][117][118]。ウミサソリ類によく似たカスマタスピス類に関しては、カブトガニ類とウミサソリ類に対して多系統群、もしくはそのいずれに内包されるなどの説はあったが、単系統群で、ウミサソリとクモガタ類に近縁である説の方が後に広く認められる[31][114][115][116][43][117]。
- ウミグモ類:
- 鋏角類の中でウミグモ類は真鋏角類より早期に分岐した説は形態学・分子系統学の両方に最も広く認められる[5][6][80]。しかし、基盤的な鋏角類とされるハベリア類などとの系統関係(どっちの方が基盤的なのか)ははっきりしておらず[1]、2000年代の分子系統解析ではウミグモ類はクモガタ類に含まれ[105][119]、もしくは残り全ての現生節足動物より早期に分岐した群とされる[120][121][27][5]。特に後者の場合、真鋏角類は大顎類と幹性類 Cormogonida をなし、鋏角類(鋏角亜門)は多系統群として解体され、代わりにウミグモ類と真鋏角類を亜門(ウミグモ亜門、真鋏角亜門)へと昇格させる見方もある[122]。ウミグモ類自体の単系統性は確実であるが、その内部系統は分子系統解析によって再編成されつつある[123][124][125]。→詳細は「ウミグモ綱 § 系統関係」を参照
2010-2020年代にかけて、真鋏角類の単系統性は確実であり、ウミグモ類と単系統群の鋏角類になる通説も再び広い支持を得られている。真鋏角類の中でカブトガニ類は基盤的で、ウミサソリ類は一般にクモガタ類の近縁と見なされる。蛛肺類以外のクモガタ類の系統関係は、未だに議論の余地がある[5][6][51][80]。
2010年代時点では、11万種以上の鋏角類が記載される[5]。百万種に及ぶ六脚類ほどではないが、節足動物の中で鋏角類は2番目に種数の多い亜門である[4]。ウミグモ類は約1300種以上、カブトガニ類は80種以上(現生種は4種のみ)、カスマタスピス類は10種以上、ウミサソリ類は250種以上が知られる[126]。10万種以上を含んだクモガタ類の中でダニとクモは大半を占め[32]、それぞれ5万5000種以上と4万3000種以上が知られる[5]。それに次いて多様なクモガタ類はザトウムシ(約6500種)・カニムシ(約3400種)・サソリ(約2000種)・ヒヨケムシ(約1100種)が挙げられており、残りのクモガタ類は種数が少なく、多くても数百種程度である[5]。
- 鋏角亜門 Chelicerata - 鋏角類。第1体節の付属肢は祖先形質としてはさみ型の鋏角/鋏肢[5][3]。
- †ハベリア目 Habeliida - ハベリア類。基盤的な鋏角類/真鋏角類。前述参照[38]。
- †モリソニア目 Mollisoniida - モリソニア類。基盤的な鋏角類/真鋏角類。前述参照[1]。
- ウミグモ綱(皆脚綱) Pycnogonida - ウミグモ類。体節は円筒状、吻と担卵肢をもつ[3]。
- 真鋏角類 Euchelicerata - 第8-13体節(後体第2-7節)の付属肢は祖先形質として蓋板状[31][5][3]。
- 節口綱(腿口綱) Merostomata(クモガタ類を除いた側系統群とされる)- 節口類。背甲は付属肢の基部以降を覆うほど発達し、脚の基節に顎基をもつ。
- カブトガニ目(剣尾目) Xiphosura - †ハラフシカブトガニ類 Synziphosurina を含む場合、次の全ての分類群を除いた側系統群とされる[31][114][115][116][117][118])
- †カスマタスピス目 Chasmataspidida
- †ウミサソリ目(広翼目) Eurypterida
- クモガタ綱 (蛛形綱、クモ綱) Arachnida - クモガタ類。背甲は付属肢を覆わず、発達した顎基と前胃を欠き、口は前腹側方向に開く[112][3]。
- ダニ亜綱 Acari - ダニ類。上唇、鋏角と触肢は顎体部(gnathosoma)をなす[3][12]。
- コヨリムシ目 Palpigradi
- †ムカシザトウムシ目 Phalangiotarbi
- ザトウムシ目 Opiliones
- クツコムシ目 Ricinulei
- ヒヨケムシ目 Solifugae
- カニムシ目 Pseudoscorpiones
- 蛛肺類 Arachnopulmonata - 後体第2-7節の範囲に書肺をもつ。
- サソリ目 Scorpiones
- Pantetrapulmonata - 後体第2-3節(第8-9体節)のみに書肺をもち、生殖口蓋は幅広く、鋏角は折りたたみナイフ状[123][119]。
- †ワレイタムシ目 Trigonotarbida
- 四肺類 Tetrapulmonata
- Serikodiastida - 後体第4-5体節に出糸器官をもち、鋏角の牙は剛毛を欠く[119]。
- †ウララネイダ目 Uraraneida
- クモ目(真正クモ目) Araneae
- Schizotarsata - 脚の跗節は3節以上の跗小節に分かれる[112][119] 。
- Serikodiastida - 後体第4-5体節に出糸器官をもち、鋏角の牙は剛毛を欠く[119]。
- 節口綱(腿口綱) Merostomata(クモガタ類を除いた側系統群とされる)- 節口類。背甲は付属肢の基部以降を覆うほど発達し、脚の基節に顎基をもつ。
進化史
[編集]


鋏角類は大顎類の節足動物と同様に古生代カンブリア紀(約5億年前)に起源していると思われ、それを直接的に示唆する化石証拠としてカンブリア紀の化石ウミグモ類[127]、および基盤的な鋏角類と思われるカンブリア紀の絶滅群ハベリア類とモリソニア類の存在が挙げられる(前述参照)[38][1]。節口類のウミサソリ類・カスマタスピス類・カブトガニ類はいずれも最古の化石記録はオルドビス紀(約4億8830万-4億4370万年前)まで遡れる[126]。既知最古のクモガタ類はシルル紀(約4億4370万-4億1600万年前)のものだが、前述の系統仮説と分子時計的証拠にあわせると、クモガタ類の起源はオルドビス紀に当たると考えられる[100][128][25]。鋏角類は水棲性を祖先形質とし、その中でクモガタ類が陸棲化した説は広く認められるが、これはクモガタ類と節口類の系統仮説の違い(特にクモガタ類の単系統性の賛否)によって解釈が変わる[51][39]。
鋏角類は古生代の初期に出現し、常に節足動物相の一翼を担ってきた。特に古生代前期には非常に繁栄し、ウミサソリ類はシルル紀にほぼ食物連鎖の頂点に位置し、史上最大の節足動物の一つになっている。また生物の陸上進出が始まったときにも早い時期に陸生種を出し、肉食者としての地位を築いた。しかし、多くは次第に衰退し、現在ではクモ類・ダニ類以外は種数もごく少なく、生きている化石的なものが多い。[要出典]
この理由として、一つには鋏角類の前体にある付属肢はほとんどが歩脚となり、口器として独立した付属肢は1対の鋏角しかなく、大顎類のように複雑で多様な口器を発達させる余地がなかったことが挙げられる。このため、その歴史の早期には強力な肉食者として存在したが、それ以降に食性についての幅広い適応を行うことができなかった。また、陸上種に関しては呼吸器としての書肺をもっていたが、そのために気管の発達が遅れ、そのために運動能力において後れを取ったとの説もある。[要出典]
しかしクモ類とダニ類は例外的で、多様化して大きな発展を遂げた。前者は糸と、それによる網の活用で広範囲な昆虫を捕食する能力を発達させたこと、後者では小さな体を利してニッチを拡大し、食性の幅を広げたことによると思われる。[要出典]



なお、鋏角類の口器は上述のように言われるほど単純で非効率的でなく、特に基盤的な真鋏角類である節口類(カブトガニ類とウミサソリ類)においては7対(鋏角+5対の脚の顎基+唇様肢/下層板)ほど多くの付属肢構造が口器に参加し、大顎類に劣らないほど複雑な咀嚼機構をなしていた[10]。クモガタ類に見られる単調な口器は祖先的でなく、むしろ陸上生活に向かって節口類の複雑な口器から高度に特化した結果と言うべきことも指摘される[10]。クモガタ類における顎基と複眼の退化は、それぞれ陸上環境への適応(顎基は水棲性の節足動物のみに見られる)と各系統群における視力以外の感覚器(例えばクモの聴毛・サソリの櫛状器・ヒヨケムシのラケット器官など)への依存に大きく関与していると考えられる[25]。
脚注
[編集]注釈
[編集]- ^
- 左上:Ammothea hilgendorfi(ウミグモ類)
- 右上:アメリカカブトガニ(節口類:カブトガニ類)
- 左中:ユーリプテルス(節口類:ウミサソリ類)
- 右中:イワオニグモ(クモガタ類:クモ類)
- 左下:Buthus occitanus(クモガタ類:サソリ類)
- 右下:Trombidium holosericeum (クモガタ類:ダニ類)
- ^ O:単眼、緑:前体、黄(1-7):中体(前腹部)、ピンク(M1-M5):終体(後腹部)、T:尾節、A:肛門、C:鋏角、P:触肢、Co・Fe・Ti・Ta:歩脚
- ^ ただしウミグモ類の場合、体は通常では頭部・胴部・腹部の3部として区別される(詳細は該当項目を参照)。サソリなどの場合、後体はさらに中体と終体に区別される。
- ^ a b 摂食用の鋏角(第1体節由来、大顎類の第3-5体節由来の顎に相似)、
カブトガニ類とウミサソリ類の顎基(脚の付け根の突起物であり、独立した付属肢ではない)、
ウデムシ・サソリモドキ・ヤイトムシの感覚用の第1脚(第3体節由来、大顎類の第1-2体節由来の触角に相似)、
などが挙げられる。 - ^ ヒヨケムシ・ヤイトムシ・コヨリムシ・ザトウムシ(痕跡的)・一部のダニ・ウミグモなど。
- ^ 一部のカブトガニ類は1対の中眼の間にさらに1つの中眼をもち、あわせて3つの単眼となる。
- ^ ヒヨケムシ・ダイオウウミサソリ科のウミサソリ・一部のクモ(トタテグモ下目・アシナガグモ科のオス・アリグモ属のオスなど)。
- ^ a b 四肺類(クモ・コスリイムシ・ウデムシ・サソリモドキ・ヤイトムシなど)。
- ^ クツコムシ・ヒヨケムシ・カニムシ・ワレイタムシ・クモ・コスリイムシ・ウデムシ・サソリモドキ・ヤイトムシなど
- ^ 歩脚状(ただし直後の脚から顕著に区別できる):ウミグモ・ダニ・コヨリムシ・ムカシザトウムシ・ザトウムシ・ヒヨケムシ・ワレイタムシ・クモ・コスリイムシ
鎌状(亜鋏状):ザトウムシ(一部)・ウデムシ・ヤイトムシ
鋏状:クツコムシ・カニムシ・サソリ・サソリモドキ - ^ カブトガニ類の最終の脚の櫂状器(flabellum)は外肢由来とも考えられる。
- ^ 多くのウミグモ(鋏肢・触肢・担卵肢は科によってあったり欠けたりする)と一部のダニ(第3-4脚を欠く分類群はいくつかある)。
- ^ サソリとウミサソリ類(前7/8節と後6/5節)、カスマタスピス類(前4節と後9節)、多くのハラフシカブトガニ類(前7/8節と後3節)。
- ^ ワレイタムシ・ウデムシ・サソリモドキ・ヤイトムシ・クツコムシ・コヨリムシなど。
- ^ 剣状(棘状):カブトガニ類、ほとんどのウミサソリ類、ほとんどのカスマタスピス類、パレオイソプス(ウミグモ)
へら状(平板状):プテリゴトゥス上科のウミサソリ類、一部のカスマタスピス類
鞭状(鞭状体):コヨリムシ、サソリモドキ、ウララネイダ類、キメララクネ(基盤的なクモ)、Flagellopantopus(ウミグモ) 鉤状(毒針):サソリ - ^ 例えばサソリの場合はこの区別方法に応じず、後体前8節を中体(=前腹部)、後5節を終体(=後腹部)と呼ばれるのが一般的であり、クモ・カニムシ・ザトウムシ・ダニなどのクモガタ類の後体は顕著な体節分化が見当たらず、一般的に中体と終体(前腹部と後腹部)の区別がなされていない。
- ^ ただし基盤的な鋏角類とされるハベリア類とモリソニア類は、それ以降の体節にも付属肢をもつ。
- ^ a b サソリ・ワレイタムシ・クモ・ウデムシ・サソリモドキ・ヤイトムシなど。
- ^ ただしサソリの前体腹面の腹板に関しては後体第1節の付属肢に由来という説がある。
- ^ ダニ(一部を除く)・ザトウムシ・カニムシ・ヒヨケムシ・クツコムシなど。
- ^ アシナガダニの気門は背側に開口し、ヒヨケムシの後体第5節の気門は中央で単一の開口になる場合がある。
- ^ I:鋏角の神経、II:触肢の神経、III-VI:脚の神経、cb:脳/食道上神経塊、n.o.l.:側眼の神経、n.o.m.:中眼の神経、op.n.:後体の神経、s.o.:食道下神経塊
- ^ br:脳/食道上神経塊、ce:後体の神経、le:側眼、me:中眼、nch:鋏角の神経、np1-np4:第1-4脚の神経、npd:触肢の神経、seg:食道下神経塊
出典
[編集]- ^ a b c d e f g h i Jean-Bernard Caron; Aria, Cédric (2019-09-11). “A middle Cambrian arthropod with chelicerae and proto-book gills” (英語). Nature: 1–4. doi:10.1038/s41586-019-1525-4. ISSN 1476-4687.
- ^ a b Heymons, R. (1901年1月1日). “Die Entwicklungsgeschichte der Scolopender” (ドイツ語). www.schweizerbart.de. 2020年4月22日閲覧。
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay Dunlop, Jason A.; Lamsdell, James C. (2017-05-01). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395–418. doi:10.1016/j.asd.2016.05.002. ISSN 1467-8039.
- ^ a b Giribet, Gonzalo; Edgecombe, Gregory (2013-10-01). Arthropod Biology and Evolution. Molecules, Development, Morphology. pp. 17–40
- ^ a b c d e f g h i j k l m n o p q r s t Dunlop, Jason; Borner, Janus; Burmester, Thorsten (2014-01-29). Wägele, J. Wolfgang; Bartolomaeus, Thomas. eds. Phylogeny of the Chelicerates: Morphological and molecular evidence. Berlin, Boston: DE GRUYTER. doi:10.1515/9783110277524.399. ISBN 978-3-11-027752-4
- ^ a b c d e f g Giribet, Gonzalo; Edgecombe, Gregory D. (2019-06-17). “The Phylogeny and Evolutionary History of Arthropods”. Current Biology 29 (12): R592–R602. doi:10.1016/j.cub.2019.04.057. ISSN 0960-9822.
- ^ “Chelicera | Definition of Chelicera by Oxford Dictionary on Lexico.com also meaning of Chelicera” (英語). Lexico Dictionaries | English. 2020年12月1日閲覧。
- ^ a b c d Bird, Tharina; Wharton, Robert (Robert Alan); Prendini, Lorenzo (2015-06-22) (英語). Cheliceral morphology in Solifugae (Arachnida) : primary homology, terminology, and character survey. (Bulletin of the American Museum of Natural History, no. 394).
- ^ a b c Fusco, Giuseppe; Minelli, Alessandro (2013). Minelli, Alessandro; Boxshall, Geoffrey; Fusco, Giuseppe. eds (英語). Arthropod Segmentation and Tagmosis. Berlin, Heidelberg: Springer. pp. 197-221. doi:10.1007/978-3-642-36160-9_9. ISBN 978-3-642-36160-9
- ^ a b c d e f g Haug, Carolin (2020-08-13). “The evolution of feeding within Euchelicerata: data from the fossil groups Eurypterida and Trigonotarbida illustrate possible evolutionary pathways” (英語). PeerJ 8: e9696. doi:10.7717/peerj.9696. ISSN 2167-8359.
- ^ a b c Dunlop, Jason A.; Penney, David (2012) (英語). Fossil Arachnids. Siri Scientific Press. ISBN 978-0-9567795-4-0
- ^ a b Bolton, Samuel J. (2022-02-25). “Proteonematalycus wagneri Kethley reveals where the opisthosoma begins in acariform mites” (英語). PLOS ONE 17 (2): e0264358. doi:10.1371/journal.pone.0264358. ISSN 1932-6203. PMC 8880937. PMID 35213630.
- ^ 日本大百科全書(ニッポニカ). “クモとは”. コトバンク. 2022年4月13日閲覧。
- ^ a b c Telford, Maximilian J.; Thomas, Richard H. (1998-09-01). “Expression of homeobox genes shows chelicerate arthropods retain their deutocerebral segment” (英語). Proceedings of the National Academy of Sciences 95 (18): 10671–10675. ISSN 0027-8424. PMID 9724762.
- ^ a b Hughes, Cynthia L.; Kaufman, Thomas C. (2002-03-01). “Exploring the myriapod body plan: expression patterns of the ten Hox genes in a centipede” (英語). Development 129 (5): 1225–1238. ISSN 0950-1991. PMID 11874918.
- ^ a b Sharma, Prashant P.; Schwager, Evelyn E.; Extavour, Cassandra G.; Giribet, Gonzalo (2012-09). “Hox gene expression in the harvestman Phalangium opilio reveals divergent patterning of the chelicerate opisthosoma: Hox genes and the chelicerate opisthosoma” (英語). Evolution & Development 14 (5): 450–463. doi:10.1111/j.1525-142X.2012.00565.x.
- ^ a b Barnett, Austen A.; Thomas, Richard H. (2013-08-30). “Posterior Hox gene reduction in an arthropod: Ultrabithorax and Abdominal-B are expressed in a single segment in the mite Archegozetes longisetosus”. EvoDevo 4 (1): 23. doi:10.1186/2041-9139-4-23. ISSN 2041-9139. PMC 3766265. PMID 23991696.
- ^ a b Sharma, Prashant P.; Santiago, Marc A.; González-Santillán, Edmundo; Monod, Lionel; Wheeler, Ward C. (2015-11). “Evidence of duplicated Hox genes in the most recent common ancestor of extant scorpions”. Evolution & Development 17 (6): 347–355. doi:10.1111/ede.12166. ISSN 1525-142X. PMID 26492826.
- ^ a b Schwager, Evelyn E.; Sharma, Prashant P.; Clarke, Thomas; Leite, Daniel J.; Wierschin, Torsten; Pechmann, Matthias; Akiyama-Oda, Yasuko; Esposito, Lauren et al. (2017-07-31). “The house spider genome reveals an ancient whole-genome duplication during arachnid evolution”. BMC Biology 15 (1): 62. doi:10.1186/s12915-017-0399-x. ISSN 1741-7007. PMC 5535294. PMID 28756775.
- ^ a b c d Ortega-Hernández, Javier; Janssen, Ralf; Budd, Graham E. (2017-05-01). “Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective” (英語). Arthropod Structure & Development 46 (3): 354–379. doi:10.1016/j.asd.2016.10.011. ISSN 1467-8039.
- ^ a b “Larval development and morphogenesis of the sea spider Pycnogonum litorale (Ström, 1762) and the tagmosis of the body of Pantopoda” (英語). Arthropod Structure & Development 32 (4): 349–383. (2003-12-01). doi:10.1016/j.asd.2003.09.004. ISSN 1467-8039.
- ^ Sharma, Prashant; Gupta, Tripti; E Schwager, Evelyn; Wheeler, W.C.; Extavour, Cassandra (2014-01-09). “Subdivision of arthropod cap-n-collar expression domains is restricted to Mandibulata”. EvoDevo 5: 3. doi:10.1186/2041-9139-5-3.
- ^ a b c d e Miether, Sebastian T.; Dunlop, Jason A. (2016/07). “Lateral eye evolution in the arachnids”. Arachnology 17 (2): 103–119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ a b c Lehmann, Tobias; Melzer, Roland R. (2018-12). “Also looking like Limulus? – retinula axons and visual neuropils of Amblypygi (whip spiders)” (英語). Frontiers in Zoology 15 (1): 52. doi:10.1186/s12983-018-0293-6. ISSN 1742-9994. PMC 6299927. PMID 30574172.
- ^ a b c Howard, Richard J.; Puttick, Mark N.; Edgecombe, Gregory D.; Lozano-Fernandez, Jesus (2020-11-01). “Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata” (英語). Arthropod Structure & Development 59: 100997. doi:10.1016/j.asd.2020.100997. ISSN 1467-8039.
- ^ a b Bicknell, Russell D. C.; Amati, Lisa; Ortega-Hernández, Javier (2019-11-14). “New insights into the evolution of lateral compound eyes in Palaeozoic horseshoe crabs” (英語). Zoological Journal of the Linnean Society 187 (4): 1061–1077. doi:10.1093/zoolinnean/zlz065. ISSN 0024-4082.
- ^ a b Dunlop, J. A.; Arango, C. P. (2005-02-01). “Pycnogonid affinities: a review” (英語). Journal of Zoological Systematics and Evolutionary Research 43 (1). doi:10.1111/j.1439-0469.2004.00284.x/abstract. ISSN 1439-0469.
- ^ “Cheliceral segmentation”. idtools.org. 2020年12月1日閲覧。
- ^ Sutton, Mark D.; Briggs, Derek E. G.; Siveter, David J.; Siveter, Derek J.; Orr, Patrick J. (2002-06-22). “The arthropod Offacolus kingi (Chelicerata) from the Silurian of Herefordshire, England: computer based morphological reconstructions and phylogenetic affinities” (英語). Proceedings of the Royal Society of London. Series B: Biological Sciences 269 (1497): 1195–1203. doi:10.1098/rspb.2002.1986. ISSN 0962-8452.
- ^ a b Briggs, Derek E. G.; Siveter, Derek J.; Siveter, David J.; Sutton, Mark D.; Garwood, Russell J.; Legg, David (2012-09-25). “Silurian horseshoe crab illuminates the evolution of arthropod limbs” (英語). Proceedings of the National Academy of Sciences 109 (39): 15702–15705. doi:10.1073/pnas.1205875109. ISSN 0027-8424. PMID 22967511.
- ^ a b c d e f g h i j k l Lamsdell, James C. (2013-01-18). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura” (英語). Zoological Journal of the Linnean Society 167 (1): 1–27. doi:10.1111/j.1096-3642.2012.00874.x. ISSN 0024-4082.
- ^ a b c d e f Dunlop, Jason A. (2019-01-01). “Miniaturisation in Chelicerata” (英語). Arthropod Structure & Development 48: 20–34. doi:10.1016/j.asd.2018.10.002. ISSN 1467-8039.
- ^ a b c Dunlop, Jason (1997) (英語). The origins of tetrapulmonate book lungs and their significance for chelicerate phylogeny.
- ^ a b c Størmer, Leif (1944) (English). On the relationships and phylogeny of fossil and recent Arachnomorpha: a comparative study on Arachnida, Xiphosura, Eurypterida, Trilobita, and other fossil Arthropoda. Oslo: Jacob Dybwad. OCLC 961296639
- ^ Di, Zhiyong; Edgecombe, Gregory D.; Sharma, Prashant P. (2018-05-21). “Homeosis in a scorpion supports a telopodal origin of pectines and components of the book lungs”. BMC Evolutionary Biology 18 (1): 73. doi:10.1186/s12862-018-1188-z. ISSN 1471-2148. PMC 5963125. PMID 29783957.
- ^ a b Shultz, Jeffrey W. (1993-08-01). “Muscular anatomy of the giant whipscorpion Mastigoproctus giganteus (Lucas) (Arachnida: Uropygi) and its evolutionary significance” (英語). Zoological Journal of the Linnean Society 108 (4): 335–365. doi:10.1111/j.1096-3642.1993.tb00302.x. ISSN 0024-4082.
- ^ a b Shultz, Jeffrey W. (1999-05-01). “Muscular anatomy of a whipspider, Phrynus longipes (Pocock) (Arachnida: Amblypygi), and its evolutionary significance” (英語). Zoological Journal of the Linnean Society 126 (1): 81–116. doi:10.1111/j.1096-3642.1999.tb00608.x. ISSN 0024-4082.
- ^ a b c d e f Aria, Cédric; Caron, Jean-Bernard (2017-12). “Mandibulate convergence in an armoured Cambrian stem chelicerate” (英語). BMC Evolutionary Biology 17 (1). doi:10.1186/s12862-017-1088-7. ISSN 1471-2148. PMC 5738823. PMID 29262772.
- ^ a b c d e f Lamsdell, James C.; McCoy, Victoria E.; Perron-Feller, Opal A.; Hopkins, Melanie J. (2020-09-10). “Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion” (English). Current Biology 0 (0). doi:10.1016/j.cub.2020.08.034. ISSN 0960-9822. PMID 32916114.
- ^ Woods, H. Arthur; Lane, Steven J.; Shishido, Caitlin; Tobalske, Bret W.; Arango, Claudia P.; Moran, Amy L. (2017-07-10). “Respiratory gut peristalsis by sea spiders” (English). Current Biology 27 (13): R638–R639. doi:10.1016/j.cub.2017.05.062. ISSN 0960-9822. PMID 28697358.
- ^ a b c BAIN, BONNIE A.; GOVEDICH, FREDRIC R. (2004-12-01). “Courtship and mating behavior in the Pycnogonida (Chelicerata: Class Pycnogonida): a summary”. Invertebrate Reproduction & Development 46 (1): 63–79. doi:10.1080/07924259.2004.9652607. ISSN 0792-4259.
- ^ Braddy, SIMON J.; Dunlop, JASON A. (1997-08-01). “The functional morphology of mating in the Silurian eurypterid,Baltoeurypterus tetragonophthalmus(Fischer, 1839)” (英語). Zoological Journal of the Linnean Society 120 (4): 435–461. doi:10.1006/zjls.1997.0093. ISSN 0024-4082.
- ^ a b Lamsdell, James C.; Gunderson, Gerald O.; Meyer, Ronald C. (2019-01-08). “A common arthropod from the Late Ordovician Big Hill Lagerstätte (Michigan) reveals an unexpected ecological diversity within Chasmataspidida”. BMC Evolutionary Biology 19 (1): 8. doi:10.1186/s12862-018-1329-4. ISSN 1471-2148. PMC 6325806. PMID 30621579.
- ^ Talarico, G.; García Hernández, L. F.; Michalik, P. (2008-09-01). “The male genital system of the New World Ricinulei (Arachnida): Ultrastructure of spermatozoa and spermiogenesis with special emphasis on its phylogenetic implications” (英語). Arthropod Structure & Development 37 (5): 396–409. doi:10.1016/j.asd.2008.01.006. ISSN 1467-8039.
- ^ a b c Smarandache-Wellmann, Carmen Ramona (10 24, 2016). “Arthropod neurons and nervous system”. Current biology: CB 26 (20): R960–R965. doi:10.1016/j.cub.2016.07.063. ISSN 1879-0445. PMID 27780069.
- ^ a b Harzsch, Steffen; Wildt, Miriam; Battelle, Barbara; Waloszek, Dieter (2005-07-01). “Immunohistochemical localization of neurotransmitters in the nervous system of larval Limulus polyphemus (Chelicerata, Xiphosura): evidence for a conserved protocerebral architecture in Euarthropoda” (英語). Arthropod Structure & Development 34 (3): 327–342. doi:10.1016/j.asd.2005.01.006. ISSN 1467-8039.
- ^ Park, Yong-Ki; Gu, Hye-Yoon; Kwon, Hyun-Jung; Kim, Hoon; Moon, Myung-Jin (2018-03-30). “Fine Structure of the Neuroganglia in the Central Nervous System of the Harvestman Leiobunum japonicum (Arachnida: Opiliones)” (英語). Applied Microscopy 48 (1): 17–26. doi:10.9729/AM.2018.48.1.17. ISSN 2287-5123.
- ^ Ryan S. Davis (October 2010). "Eriophyid Mites: bud, blister, gall, and rust mites" (PDF). Utah Pests Fact Sheet. Utah State University Extension and Utah Plant Pest Diagnostic Laboratory. Retrieved January 26, 2011.
- ^ a b Tetlie, O. Erik (2007-09-03). “Distribution and dispersal history of Eurypterida (Chelicerata)” (英語). Palaeogeography, Palaeoclimatology, Palaeoecology 252 (3): 557–574. doi:10.1016/j.palaeo.2007.05.011. ISSN 0031-0182.
- ^ Braddy, Simon J; Poschmann, Markus; Tetlie, O. Erik (2008-02-23). “Giant claw reveals the largest ever arthropod”. Biology Letters 4 (1): 106–109. doi:10.1098/rsbl.2007.0491. ISSN 1744-9561. PMC 2412931. PMID 18029297.
- ^ a b c d e f g Howard, Richard J.; Edgecombe, Gregory D.; Legg, David A.; Pisani, Davide; Lozano-Fernandez, Jesus (2019-03-01). “Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks” (英語). Organisms Diversity & Evolution 19 (1): 71–86. doi:10.1007/s13127-019-00390-7. ISSN 1618-1077.
- ^ Whyte, Martin A. (2005-12). “A gigantic fossil arthropod trackway” (英語). Nature 438 (7068): 576–576. doi:10.1038/438576a. ISSN 1476-4687.
- ^ 島野智之『なぜダニ類はクモガタ類の中で最も種数が多いのか?』日本動物分類学会、2018年。doi:10.19004/taxa.44.0_4。2020年4月28日閲覧。
- ^ Jackson, Robert R.; Pollard, Simon D.; Nelson, Ximena J.; Edwards, G. B.; Barrion, Alberto T.. “Jumping spiders (Araneae: Salticidae) that feed on nectar” (英語). Journal of Zoology 255 (1): 25–29. ISSN 0952-8369.
- ^ 伊賀タウン情報YOU編集部 (2019年8月4日). “蜜吸うクモ 機関紙で論文発表 名張の岡田さん | 【伊賀タウン情報 YOU】”. 2020年4月27日閲覧。
- ^ Meehan, Christopher J.; Olson, Eric J.; Reudink, Matthew W.; Kyser, T. Kurt; Curry, Robert L. (2009-10-13). “Herbivory in a spider through exploitation of an ant–plant mutualism” (English). Current Biology 19 (19): R892–R893. doi:10.1016/j.cub.2009.08.049. ISSN 0960-9822. PMID 19825348.
- ^ Botton, M. L. (1984-08-01). “Diet and food preferences of the adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA” (英語). Marine Biology 81 (2): 199–207. doi:10.1007/BF00393118. ISSN 1432-1793.
- ^ a b Dietz, Lars; Dömel, Jana S.; Leese, Florian; Lehmann, Tobias; Melzer, Roland R. (2018-03-15). “Feeding ecology in sea spiders (Arthropoda: Pycnogonida): what do we know?”. Frontiers in Zoology 15 (1): 7. doi:10.1186/s12983-018-0250-4. ISSN 1742-9994. PMC 5856303. PMID 29568315.
- ^ Shultz, Jeffrey W. (2000-04-01). “Skeletomuscular anatomy of the harvestman Leiobunum aldrichi (Weed, 1893) (Arachnida: Opiliones: Palpatores) and its evolutionary significance” (英語). Zoological Journal of the Linnean Society 128 (4): 401–438. doi:10.1111/j.1096-3642.2000.tb01522.x. ISSN 0024-4082.
- ^ “Atlantic Horseshoe Crab”. Digestive systems. 2020年4月27日閲覧。
- ^ Brenneis, Georg; Bogomolova, Ekaterina V.; Arango, Claudia P.; Krapp, Franz (2017-02-07). “From egg to “no-body”: an overview and revision of developmental pathways in the ancient arthropod lineage Pycnogonida”. Frontiers in Zoology 14 (1): 6. doi:10.1186/s12983-017-0192-2. ISSN 1742-9994. PMC 5297176. PMID 28191025.
- ^ Alexeeva, Nina; Tamberg, Yuta; Shunatova, Natalia (2018-05-01). “Postembryonic development of pycnogonids: A deeper look inside” (英語). Arthropod Structure & Development 47 (3): 299–317. doi:10.1016/j.asd.2018.03.002. ISSN 1467-8039.
- ^ Chapin, Kenneth J.; Hebets, Eileen A.. “The behavioral ecology of amblypygids” (英語). Journal of Arachnology 44 (1): 1–14. ISSN 0161-8202.
- ^ “Opiliones - an overview | ScienceDirect Topics”. www.sciencedirect.com. 2020年4月28日閲覧。
- ^ a b c 1911 Encyclopædia Britannica, Volume 2: Arthropoda by Edwin Ray Lankester 2022年4月13日閲覧。
- ^ a b Savigny, J.C. 1816. Mémoires sur les animaux sans vertèbres. Paris: l’Imprimerie de Crapelet, 239 pp.
- ^ a b Blanchard, É. 1847. Observations sur l’organisation d’un type de la classe des Arachnides, le genre Galéode (Galeodes Latr.). Annales des Sciences Naturelles, Zoologie (3) 8: 227–238, plate 6.
- ^ a b c Latreille, P.A. 1829. Les crustacés, les arachnides et partie des insectes. In G. Cuvier (editor), Le règne animal distribué d’après son organisation, pour servir de base a l’histoire natúrelle des animaux et d’introduction l’anatomie comparée. Avec figures dessinées d’après nature. 2nd ed., Vol. 4: 73–125. Paris: Chez Déterville.
- ^ a b Viallanes, H. 1892. Études histologiques et organologiques sur les centres nerveux et les organes des sens des animaux articulés (6th mémoire). Annales des Sciences Naturelles, Zoologie 14: 74 pp., 5 plates.
- ^ a b Woodward, Henry (1866). A monograph of the British fossil Crustacea, belonging to the order Merostomata.. London,: Printed for the Palæontographical Society,. doi:10.5962/bhl.title.53733
- ^ a b c d Stürmer, Wilhelm; Bergström, Jan (1978-06-01). “The arthropod Cheloniellon from the devonian hunsrück shale” (英語). Paläontologische Zeitschrift 52 (1): 57–81. doi:10.1007/BF03006730.
- ^ a b c d Stürmer, Wilhelm; Bergström, Jan (1981-12-01). “Weinbergina, a xiphosuran arthropod from the devonian hunsrück slate” (ドイツ語). Paläontologische Zeitschrift 55 (3): 237–255. doi:10.1007/BF02988142.
- ^ Damen, Wim G. M. (2002-03-01). “Parasegmental organization of the spider embryo implies that the parasegment is an evolutionary conserved entity in arthropod embryogenesis” (英語). Development 129 (5): 1239–1250. ISSN 0950-1991. PMID 11874919.
- ^ Jager, Muriel; Murienne, Jérôme; Clabaut, Céline; Deutsch, Jean; Guyader, Hervé Le; Manuel, Michaël (2006-05). “Homology of arthropod anterior appendages revealed by Hox gene expression in a sea spider” (英語). Nature 441 (7092): 506–508. doi:10.1038/nature04591. ISSN 1476-4687.
- ^ a b Manuel, Michaël; Jager, Muriel; Murienne, Jérôme; Clabaut, Céline; Guyader, Hervé Le (2006-07-13). “Hox genes in sea spiders (Pycnogonida) and the homology of arthropod head segments” (英語). Development Genes and Evolution 216 (7-8): 481–491. doi:10.1007/s00427-006-0095-2. ISSN 0949-944X.
- ^ Brenneis, Georg; Ungerer, Petra; Scholtz, Gerhard (2008-11). “The chelifores of sea spiders (Arthropoda, Pycnogonida) are the appendages of the deutocerebral segment”. Evolution & Development 10 (6): 717–724. doi:10.1111/j.1525-142X.2008.00285.x. ISSN 1525-142X. PMID 19021742.
- ^ a b Mittmann, Beate; Scholtz, Gerhard (2003-02). “Development of the nervous system in the "head" of Limulus polyphemus (Chelicerata: Xiphosura): morphological evidence for a correspondence between the segments of the chelicerae and of the (first) antennae of Mandibulata”. Development Genes and Evolution 213 (1): 9–17. doi:10.1007/s00427-002-0285-5. ISSN 0949-944X. PMID 12590348.
- ^ Maxmen, Amy; Browne, William E.; Martindale, Mark Q.; Giribet, Gonzalo (2005-10). “Neuroanatomy of sea spiders implies an appendicular origin of the protocerebral segment” (英語). Nature 437 (7062): 1144–1148. doi:10.1038/nature03984. ISSN 1476-4687.
- ^ a b Regier, Jerome C.; Shultz, Jeffrey W.; Zwick, Andreas; Hussey, April; Ball, Bernard; Wetzer, Regina; Martin, Joel W.; Cunningham, Clifford W. (2010-02). “Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences” (英語). Nature 463 (7284): 1079–1083. doi:10.1038/nature08742. ISSN 0028-0836.
- ^ a b c d Edgecombe, Gregory D. (2020-07-31). “Arthropod Origins: Integrating Paleontological and Molecular Evidence”. Annual Review of Ecology, Evolution, and Systematics. doi:10.1146/annurev-ecolsys-011720-124437. ISSN 1543-592X.
- ^ a b Tanaka, Gengo; Hou, Xianguang; Ma, Xiaoya; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (17 October 2013). “Chelicerate neural ground pattern in a Cambrian great appendage arthropod”. Nature 502 (7471): 364–367. doi:10.1038/nature12520. PMID 24132294.
- ^ Liu, Yu; Ortega-Hernández, Javier; Zhai, Dayou; Hou, Xianguang (2020-06-25). “A Reduced Labrum in a Cambrian Great-Appendage Euarthropod” (English). Current Biology 0 (0). doi:10.1016/j.cub.2020.05.085. ISSN 0960-9822. PMID 32589912.
- ^ a b c Chen, Junyuan; Waloszek, Dieter; Maas, Andreas (2004). “A new ‘great-appendage’ arthropod from the Lower Cambrian of China and homology of chelicerate chelicerae and raptorial antero-ventral appendages” (英語). Lethaia 37 (1): 3–20. doi:10.1080/00241160410004764. ISSN 1502-3931.
- ^ a b c Trevor Cotton, S.J. Braddy. (2004-1). The phylogeny of arachnomorph arthropods and the origin of the Chelicerata. Transactions of the Royal Society of Edinburgh Earth Sciences 94 (3): 169-193.
- ^ a b c Dunlop, Jason A. (2005-11). New ideas about the euchelicerate stem-lineage. Acta Zoologica Bulgarica.
- ^ a b c Haug, Joachim T.; Waloszek, Dieter; Maas, Andreas; Liu, Yu; Haug, Carolin (2012-03). “Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian” (英語). Palaeontology 55 (2): 369–399. doi:10.1111/j.1475-4983.2011.01124.x.
- ^ a b Ortega-Hernández, Javier; Lerosey-Aubril, Rudy; Losso, Sarah R.; Weaver, James C. (2022-01-20). “Neuroanatomy in a middle Cambrian mollisoniid and the ancestral nervous system organization of chelicerates” (英語). Nature Communications 13 (1): 410. doi:10.1038/s41467-022-28054-9. ISSN 2041-1723.
- ^ a b Legg, David A. (2014-12-01). “Sanctacaris uncata: the oldest chelicerate (Arthropoda)” (英語). Naturwissenschaften 101 (12): 1065–1073. doi:10.1007/s00114-014-1245-4. ISSN 1432-1904.
- ^ Gould, Stephen Jay; 渡辺, 政隆 (2000) (Japanese). ワンダフル・ライフ: バージェス頁岩と生物進化の物語. ISBN 978-4-15-050236-2. OCLC 676428996
- ^ a b Ortega-Hernández, J.; Legg, D. A.; Braddy, S. J. (2013). “The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda”. Cladistics 29: 15–45. doi:10.1111/j.1096-0031.2012.00413.x.
- ^ a b Scholtz, Gerhard; Edgecombe, Gregory (2005-04-27). Koenemann, Stefan; Jenner, Ronald. eds (英語). Heads, Hox and the phylogenetic position of trilobites. Crustacea and Arthropod Phylogeny. –In book: Crustacea and Arthropod Relationships. 16. CRC Press. pp. 139–165. doi:10.1201/9781420037548.ch6. ISBN 978-0-8493-3498-6
- ^ Legg, David A.; Sutton, Mark D.; Edgecombe, Gregory D. (2013-09-30). “Arthropod fossil data increase congruence of morphological and molecular phylogenies” (英語). Nature Communications 4 (1): 2485. doi:10.1038/ncomms3485. ISSN 2041-1723.
- ^ Scholtz, Gerhard; Staude, Andreas; Dunlop, Jason A. (2019-06-07). “Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinities” (英語). Nature Communications 10 (1): 1–7. doi:10.1038/s41467-019-10459-8. ISSN 2041-1723.
- ^ Chen, Jun-Yuan (2009). “The sudden appearance of diverse animal body plansduring the Cambrian explosion”. The International Journal of Developmental Biology 53 (5-6): 733–751. doi:10.1387/ijdb.072513cj. ISSN 1696-3547. PMID 19557680.
- ^ 日本産クモ類. 小野展嗣 (Dai 1-han ed.). 東海大学出版会,. (2009). ISBN 978-4-486-01791-2. OCLC 429985205
- ^ Ortega‐Hernández, Javier (2016). “Making sense of ‘lower’ and ‘upper’ stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848” (英語). Biological Reviews 91 (1): 255–273. doi:10.1111/brv.12168. ISSN 1469-185X.
- ^ Peiyun Cong; Xiaoya Ma; Xianguang Hou; Gregory D. Edgecombe; Nicholas J. Strausfeld (2014). “Brain structure resolves the segmental affinity of anomalocaridid appendages”. Nature 513 (7519): 538–42. doi:10.1038/nature13486. PMID 25043032.
- ^ a b c Sharma, Prashant P.; Kaluziak, Stefan T.; Pérez-Porro, Alicia R.; González, Vanessa L.; Hormiga, Gustavo; Wheeler, Ward C.; Giribet, Gonzalo (2014-11-01). “Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal” (英語). Molecular Biology and Evolution 31 (11): 2963–2984. doi:10.1093/molbev/msu235. ISSN 0737-4038.
- ^ a b c Ballesteros, Jesús A.; Sharma, Prashant P. (2019-11-01). “A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error” (英語). Systematic Biology 68 (6): 896–917. doi:10.1093/sysbio/syz011. ISSN 1063-5157.
- ^ a b c Lozano-Fernandez, Jesus; Tanner, Alastair R.; Giacomelli, Mattia; Carton, Robert; Vinther, Jakob; Edgecombe, Gregory D.; Pisani, Davide (2019-05-24). “Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida” (英語). Nature Communications 10 (1): 1–8. doi:10.1038/s41467-019-10244-7. ISSN 2041-1723.
- ^ Leite, Daniel J.; Baudouin-Gonzalez, Luís; Iwasaki-Yokozawa, Sawa; Lozano-Fernandez, Jesus; Turetzek, Natascha; Akiyama-Oda, Yasuko; Prpic, Nikola-Michael; Pisani, Davide et al. (09 01, 2018). “Homeobox Gene Duplication and Divergence in Arachnids”. Molecular Biology and Evolution 35 (9): 2240–2253. doi:10.1093/molbev/msy125. ISSN 1537-1719. PMC 6107062. PMID 29924328.
- ^ Gainett, Guilherme; Sharma, Prashant P. (2020-07-03). “New genomic resources and toolkits for developmental study of whip spiders (Amblypygi) provide insights into arachnid genome evolution and antenniform leg patterning” (英語). bioRxiv: 2020.07.03.185769. doi:10.1101/2020.07.03.185769.
- ^ Scholtz, Gerhard; Kamenz, Carsten (2006). “The book lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): evidence for homology and a single terrestrialisation event of a common arachnid ancestor”. Zoology (Jena, Germany) 109 (1): 2–13. doi:10.1016/j.zool.2005.06.003. ISSN 0944-2006. PMID 16386884.
- ^ 島野智之「ダニ類の高次分類体系の改訂について-高次分類群の一部の和名改称を含む-」『J日本ダニ学会誌』第27巻第2号、日本ダニ学会、2018年11月25日、51–68頁、doi:10.2300/acari.27.51、ISSN 0918-1067。
- ^ a b Masta, Susan E.; McCall, Andrew; Longhorn, Stuart J. (2010-10-01). “Rare genomic changes and mitochondrial sequences provide independent support for congruent relationships among the sea spiders (Arthropoda, Pycnogonida)” (英語). Molecular Phylogenetics and Evolution 57 (1): 59–70. doi:10.1016/j.ympev.2010.06.020. ISSN 1055-7903.
- ^ Giribet, Gonzalo; Edgecombe, Gregory D.; Wheeler, Ward C.; Babbitt, Courtney (2002). “Phylogeny and Systematic Position of Opiliones: A Combined Analysis of Chelicerate Relationships Using Morphological and Molecular Data1” (英語). Cladistics 18 (1): 5–70. doi:10.1111/j.1096-0031.2002.tb00140.x. ISSN 1096-0031.
- ^ a b Noah, Katherine E; Hao, Jiasheng; Li, Luyan; Sun, Xiaoyan; Foley, Brian; Yang, Qun; Xia, Xuhua (2020-01). “Major Revisions in Arthropod Phylogeny Through Improved Supermatrix, With Support for Two Possible Waves of Land Invasion by Chelicerates” (英語). Evolutionary Bioinformatics 16: 117693432090373. doi:10.1177/1176934320903735. ISSN 1176-9343. PMC 7003163. PMID 32076367.
- ^ Sharma, Prashant P.; Ballesteros, Jesús A.; Santibáñez-López, Carlos E. (2021-11). “What Is an “Arachnid”? Consensus, Consilience, and Confirmation Bias in the Phylogenetics of Chelicerata” (英語). Diversity 13 (11): 568. doi:10.3390/d13110568. ISSN 1424-2818.
- ^ Weygoldt, Peter (2018年). “Current views on chelicerate phylogeny—A tribute to Peter Weygoldt” (英語). 2018年11月17日閲覧。
- ^ Weygoldt, P.; Paulus, H. F. (1979). “Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata” (英語). Journal of Zoological Systematics and Evolutionary Research 17 (3): 177–200. doi:10.1111/j.1439-0469.1979.tb00699.x. ISSN 1439-0469.
- ^ Shultz, Jeffrey W. (1990). “Evolutionary Morphology and Phylogeny of Araghnida” (英語). Cladistics 6 (1): 1–38. doi:10.1111/j.1096-0031.1990.tb00523.x. ISSN 1096-0031.
- ^ a b c d e Shultz, Jeffrey W. (2007). “A phylogenetic analysis of the arachnid orders based on morphological characters” (英語). Zoological Journal of the Linnean Society 150 (2): 221–265. doi:10.1111/j.1096-3642.2007.00284.x. ISSN 1096-3642.
- ^ “[https://jglobal.jst.go.jp/detail?JGLOBAL_ID=201606004849214818 ハラフシカブトガニ亜目 | 科学技術用語情報 | J-GLOBAL 科学技術総合リンクセンター jglobal.jst.go.jp]”. 2020年8月17日閲覧。
- ^ a b c Selden, Paul A.; Lamsdell, James C.; Qi, Liu (2015). “An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China” (英語). Zoologica Scripta 44 (6): 645–652. doi:10.1111/zsc.12124. ISSN 1463-6409.
- ^ a b c Lamsdell, James C.; Briggs, Derek E. G.; Liu, Huaibao P.; Witzke, Brian J.; McKay, Robert M. (2015-10). “A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids” (英語). The Science of Nature 102 (9-10): 63. doi:10.1007/s00114-015-1312-5. ISSN 0028-1042.
- ^ a b c Lamsdell, James C. (2016-03). Zhang, Xi-Guang. ed. “Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation” (英語). Palaeontology 59 (2): 181–194. doi:10.1111/pala.12220.
- ^ a b c Bicknell, Russell D. C.; Lustri, Lorenzo; Brougham, Tom (2019-12-01). “Revision of “Bellinurus” carteri (Chelicerata: Xiphosura) from the Late Devonian of Pennsylvania, USA” (英語). Comptes Rendus Palevol 18 (8): 967–976. doi:10.1016/j.crpv.2019.08.002. ISSN 1631-0683.
- ^ a b Bicknell, Russell D. C.; Pates, Stephen (2020). “Pictorial Atlas of Fossil and Extant Horseshoe Crabs, With Focus on Xiphosurida” (English). Frontiers in Earth Science 8. doi:10.3389/feart.2020.00098. ISSN 2296-6463.
- ^ a b c d e Garwood, Russell J.; Dunlop, Jason (2014-11-13). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359.
- ^ Giribet, Gonzalo; Edgecombe, Gregory D.; Wheeler, Ward C. (2001-09). “Arthropod phylogeny based on eight molecular loci and morphology” (英語). Nature 413 (6852): 157–161. doi:10.1038/35093097. ISSN 1476-4687.
- ^ Maxmen, Amy; Browne, William E.; Martindale, Mark Q.; Giribet, Gonzalo (2005-10). “Neuroanatomy of sea spiders implies an appendicular origin of the protocerebral segment” (英語). Nature 437 (7062): 1144–1148. doi:10.1038/nature03984. ISSN 0028-0836.
- ^ Legg, David (2013-06). The impact of fossils on arthropod phylogeny.
- ^ a b Arango, Claudia P.; Wheeler, Ward C. (2007-06). “Phylogeny of the sea spiders (Arthropoda, Pycnogonida) based on direct optimization of six loci and morphology” (英語). Cladistics 23 (3): 255–293. doi:10.1111/j.1096-0031.2007.00143.x. ISSN 0748-3007.
- ^ Arabi, Juliette; Cruaud, Corinne; Couloux, Arnaud; Hassanin, Alexandre. “Studying sources of incongruence in arthropod molecular phylogenies: Sea spiders (Pycnogonida) as a case study” (英語). Comptes Rendus Biologies 333 (5): 438–453. ISSN 1631-0691.
- ^ Ballesteros, Jesús A.; Setton, Emily V. W.; López, Carlos E. Santibáñez; Arango, Claudia P.; Brenneis, Georg; Brix, Saskia; Cano-Sánchez, Esperanza; Dandouch, Merai et al. (2020-02-02). “Phylogenomic resolution of sea spider diversification through integration of multiple data classes” (英語). bioRxiv: 2020.01.31.929612. doi:10.1101/2020.01.31.929612.
- ^ a b Dunlop, J. A., Penney, D. & Jekel, D. 2020. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 20.5
- ^ Waloszek, Dieter; Dunlop, Jason A. (2002-05). “A Larval Sea Spider (Arthropoda: Pycnogonida) from the Upper Cambrian 'orsten' of Sweden, and the Phylogenetic Position of Pycnogonids” (英語). Palaeontology 45 (3): 421–446. doi:10.1111/1475-4983.00244. ISSN 0031-0239.
- ^ Lozano-Fernandez, Jesus; Tanner, Alastair R.; Puttick, Mark N.; Vinther, Jakob; Edgecombe, Gregory D.; Pisani, Davide (2020). “A Cambrian–Ordovician Terrestrialization of Arachnids” (English). Frontiers in Genetics 11. doi:10.3389/fgene.2020.00182. ISSN 1664-8021.
参考文献
[編集]- 内田亨監修『動物系統分類学(全10巻)第7巻(中A) 節足動物(IIa)』,(1966),中山書店
- 石川良輔編『節足動物の多様性と系統』,(2008),バイオディバーシティ・シリーズ6(裳華房)
- 小野展嗣編著、『日本産クモ類』、(2009)、東海大学出版会